Основи біоорганічної хімії (навчальний посібник) - Г. О. Сирова - 2018
Тема
Структура та біохімічні функції нуклеозидів, нуклеотидів і нуклеїнових кислот
Актуальність теми. Одна з найцікавіших і фундаментальних проблем, пов'язаних із синтезом білка, полягає у виявленні того, що змушує амінокислоти, які входять до складу білка, поєднуватися між собою в певній послідовності, строго визначеній для білка кожного типу. Із цим тісно пов'язане питання про те, яким чином інформація про послідовність амінокислот відтворюється в кожному новому поколінні клітин. На сьогодні відомо, що існують речовини, які містяться в хромосомах клітинних ядер, відповідають за передачу спадкових ознак (генетичної інформації) і беруть участь в управлінні біосинтезом білка. Оскільки ці речовини виділені з ядра клітини (nucleus - ядро) і мають кислотні властивості, вони називаються нуклеїновими кислотами.
Знання про будову та властивості цих найважливіших біополімерів необхідне майбутньому лікарю для розуміння сутності нормальних процесів і патології, походження низки спадкових захворювань, перспективних проблем управління деякими процесами життєдіяльності.
Загальна мета: вивчити будову і біологічні функції нуклеїнових кислот - найважливіших структурних компонентів клітини на рівні їх первинної та вторинної структури.
Конкретні цілі:
1. Аналізувати значення мононуклеотидів для будови нуклеїнових кислот і дію нуклеотидних коферментів.
2. Інтерпретувати механізми участі вітамінів у побудові коферментів, що каталізують біохімічні перетворення в організмі.
Теоретичні питання
1. Нуклеїнові кислоти - біополімери, що зберігають, передають спадкову інформацію та беруть участь у біосинтезі білка.
2. Склад і структура компонентів нуклеїнових кислот: азотистих основ, моносахаридів.
3. Нуклеозиди і нуклеотиди - продукти неповного гідролізу нуклеїнових кислот. Структура нуклеотидів.
4. Будова й значення 3', 5'-цАМФ.
5. Будова та біохімічні функції ДНК.
6. Типи РНК, їх структурна організація та біологічна роль. Відмінності в будові, локалізації і функціях ДНК і РНК.
7. Фосфорильовані похідні нуклеотидів. Значення АДФ, АТФ.
8. Участь нуклеотидів у побудові коферментів. Механізм дії НАД+.
1. Нуклеїнові кислоти (nucleus - ядро) уперше виявлені 1968 року швейцарським хіміком Мішером Ф. у ядрах клітин. Пізніше аналогічні речовини було знайдено також у протоплазмі клітин.
Нуклеїнові кислоти забезпечують зберігання і передачу спадкових ознак, беруть безпосередню участь у синтезі клітинних білків. Вони входять у структуру складних білків - нуклеопротеїдів, що містяться у всіх клітинах організму людини, тварин, рослин, бактерій, вірусів. Вміст нуклеїнових кислот у нуклеопротеїдах, крім вірусних, коливається в межах 40-65 %.
Нуклеїнові кислоти - біополімери, мономерами яких є мононуклеотиди, що гідролізуються з утворенням піримідинової або пуринової основи,
моносахаридів рибози або дезоксирибози та фосфатної кислоти. Нуклеїнові кислоти є полінуклеотидами.
2. Нуклеїнові кислоти, до складу яких входить дезоксирибоза, називаються дезоксирибонуклеїновими (ДНК). У складі рибонуклеїнових кислот (РНК) є моносахарид рибоза. Обидва моносахариди входять до складу нуклеїнових кислот у β-фуранозній формі:

Азотисті основи, що утворюють нуклеїнові кислоти, є похідними піримідину або пурину.
Основними нуклеїновими основами є 3 піримідинові (урацил, тимін, цитозин) і 2 пуринові основи (аденін, гуанін):
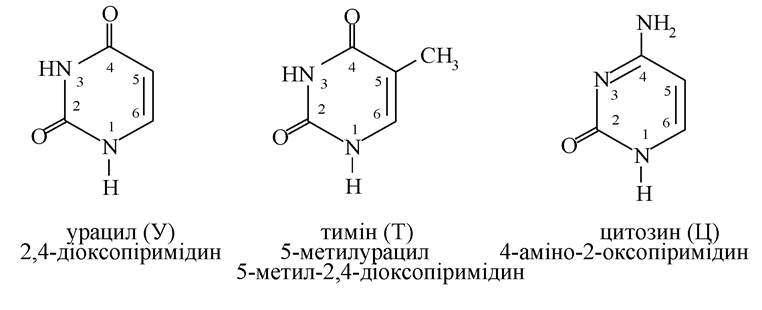
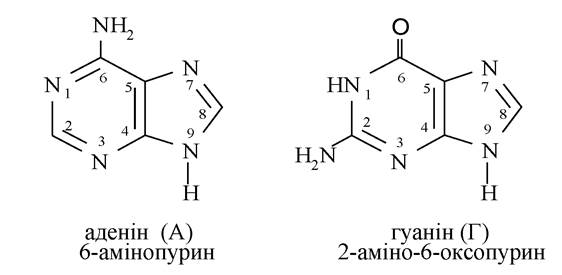
Тимін входить тільки до складу ДНК, урацил - тільки РНК, інші основи - і в ДНК, і в РНК. Оксопохідним пурину і піримідину, у зв'язку з тим, що ці сполуки виявляють NH-кислотність, характерна лактим-лактамна таутомерія:
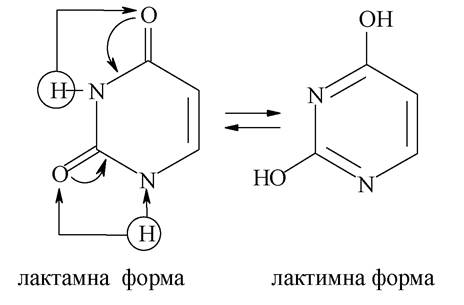
З двох форм стабільніша лактамна (оксо-) форма, тому до складу нуклеїнових кислот азотисті основи входять у лактамній формі.
3. Гідроліз нуклеїнових кислот можна показати на схемі:

Відповідно схема формування первинної структури нуклеїнової кислоти (полінуклеотиду) має зворотній напрямок.
Зв'язок між моносахаридом і основою здійснюється за участю глікозидного гідроксилу рибози або дезоксирибози і першого атома нітрогену піримідинової основи або дев'ятого атома нітрогену пуринової основи:

Цей зв'язок називається N-глікозидним зв'язком, а сполуки, які утворюються, N-глікозидами або нуклеозидами. Назви нуклеозидів утворюють від тривіальних назв основ за допомогою закінчень -идин (-ідин) для піримідинових і -озин для пуринових нуклеозидів: уридин (У), тимідин (Т), цитидін (Ц), дезоксицитидин (ДЦ), аденозин (А), дезоксиаденозин (дА), гуанозин (Г), дезоксигуанозин (дГ). Нуклеозиди стійкі до гідролізу в слабколужному середовищі, розщеплюються в кислому середовищі з утворенням вихідних речовин.
Нуклеотиди - це фосфати нуклеозидів за 5'-положенням залишку моносахариду. Залежно від структури пентози розрізняють рибонуклеотиди (мономери РНК) і дезоксирибонуклеотиди (мономери ДНК).
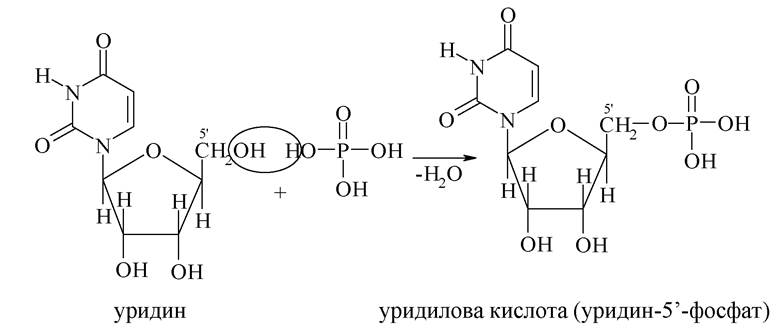
Нуклеотиди можна розглядати, з одного боку, як естери (фосфати), з іншого боку - як кислоти, оскільки містять залишок фосфатної кислоти. Тому для нуклеотидів використовують два види назв.
Найважливіші нуклеотиди, що входять до складу нуклеїнових кислот
Назва нуклеотидів |
Скорочена назва |
|
як монофосфатів* |
як кислот |
|
Аденозин-5'-фосфат |
5'-аденілова кислота |
АМФ |
Г уанозин-5'-фосфат |
5'-гуанілова кислота |
ГМФ |
Цитидин-5'-фосфат |
5'-цитидилова кислота |
ЦМФ |
Уридин-5'-фосфат |
5'-уридилова кислота |
УМФ |
Дезоксиаденозин-5'-фосфат |
Дезоксиаденілова кислота |
дАМФ |
Дезоксигуанозин-5'-фосфат |
Дезоксигуаніловаа кислота |
дГМФ |
Дезоксицитидин-5'-фосфат |
Дезоксицитидилова кислота |
дЦМФ |
Тимідин-5'-фосфат |
Тимідилова кислота |
дТМФ |
* - замість монофосфат часто пишуть просто фосфат.
4. Відомі нуклеотиди, у яких фосфорна кислота етерифікує два спиртових гідроксили моносахариду - 3' і 5'. Практично у всіх клітинах присутні два циклофосфати - циклічна 3',5'-аденілова (цАМФ) і циклічна 3',5'-гуанілова (цГМФ) кислоти.
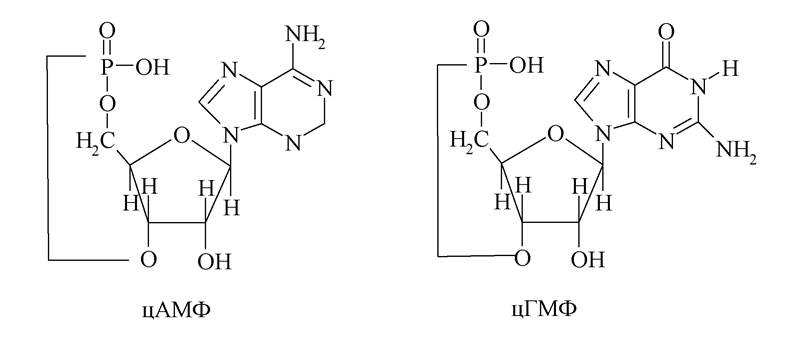
Ці нуклеотиди беруть участь у регуляції найважливіших внутрішньоклітинних процесів. Наприклад, цАМФ перетворює інертні білки на ферменти, так звані цАМФ-залежні протеїнкінази. Під дією цих ферментів відбуваються біохімічні реакції, які лежать в основі проведення нервових імпульсів та ін.
5. Утворення первинної структури НК - полінуклеотиду - відбувається внаслідок етерифікації спиртового гідроксилу при С-3' моносахариду попереднього мононуклеотиду залишком фосфорної кислоти при С-5' наступного мононуклеотиду. При цьому рибонуклеотиди утворюють РНК, дезоксирибонуклеотиди - ДНК.
Наприклад, первинну структуру ділянки ланцюга ДНК дА-дЦ-дГ-Т можна подати так (див. рис. 7).
Принцип побудови ланцюга РНК такий же, але з двома винятками: пентозним залишком у РНК є β-D-рибофураноза і в наборі гетероциклічних основ замість тиміну присутній урацил.
Нуклеотидний склад нуклеїнових кислот служить важливою характеристикою. Генетична інформація, тобто інформація про синтез певних білків, записана (закодована) у нуклеотидній послідовності ДНК. Одну амінокислоту кодує триплет нуклеотидів, який називається кодон.

Малюнок 7 - Первинну структуру ділянки ланцюга ДНК дА-дЦ-дГ-Т
Вторинна структура ДНК - просторова організація полінуклеотидних ланцюгів у її молекулі. Вторинна структура ДНК є подвійною вправозакрученою спіраллю (Дж.Уотсон і Ф.Крик, 1953 год), фіксованою вандер-ваальсовими силами, які діють уздовж спіралі, а також стабілізованою водневими зв'язками між комплементарними азотистими основами двох ниток. Діаметр спіралі становить 1,8-2,0 нм, кожен виток спіралі містить 10 пар основ. Крок спіралі дорівнює 3,4 нм. Відстань між площами основ - 0,34 нм. Полінуклеотидні ланцюги в спіралі антипаралельні, тобто в одному ланцюгу фосфодиефірні зв'язки утворюються в напрямку 3'→5', в іншому - 5'→3'.

Комплементарність ланцюгів у подвійній спіралі ДНК
У зв'язку із цим нуклеотидний склад ДНК різного походження підпорядковується закономірностям, які називаються правилами Чаргаффа:
1) кількість пуринових основ дорівнює кількості піримідинових основ: А + Г = Ц + Т;
2) кількість аденіну дорівнює кількості тиміну (А = Т), кількість гуаніну дорівнює кількості цитозину (Г = Ц);
3) кількість основ, що містять аміногрупу в положенні 4 піримідинового циклу і положенні 6 пуринового циклу, дорівнює кількості основ, що містять у цих же положеннях оксогрупу, тобто А + Ц = Г + Т.
На РНК правила Чаргаффа не поширюються.
Комплементарність ланцюгів є хімічною основою найважливішої функції ДНК - зберігання і передачі спадкових ознак.
Під час ділення клітин подвійна спіраль розкручується і поділяється на два ланцюги. На кожному із цих ланцюгів, як на матриці, відбувається біосинтез нової ДНК за принципом комплементарності. У результаті утворюються 2 нові однакові двоспіральні молекули ДНК, одна з яких залишається в батьківській клітині, а друга переходить у дочірню клітину. Цей процес називається реплікацією. Аналогічно на деспіралізованому ланцюгу ДНК у ядрі відбувається синтез молекули матричної (інформаційної) РНК, яка слугує матрицею для біосинтезу білка в цитоплазмі. Синтез мРНК є переписуванням генетичної інформації з ДНК на мРНК і називається транскрипцією.
ДНК міститься в основному в ядрах клітин, РНК переважно знаходиться в рибосомах та протоплазмі клітин. Загальна роль РНК полягає в безпосередній участі в біосинтезі білка.
6. Залежно від функцій виділяють три види РНК: транспортна РНК (тРНК), матрична РНК (мРНК) і рибосомна РНК (рРНК). Вони розрізняються за місцем розташування в клітині, складом і розмірами, а також функціями.
На частку тРНК припадає 10-20 % від усіх клітинних РНК. Основна роль тРНК - транспортування амінокислот до місця синтезу білка - у рибосоми. Макромолекула тРНК - єдиний ланцюг однотяжова) структура), який у просторі складається, утворюючи вторинну структуру, що отримала назву «листок конюшини». Для цієї структури характерна наявність чотирьох зон зі спареними основами та трьох петель з неспареними основами.
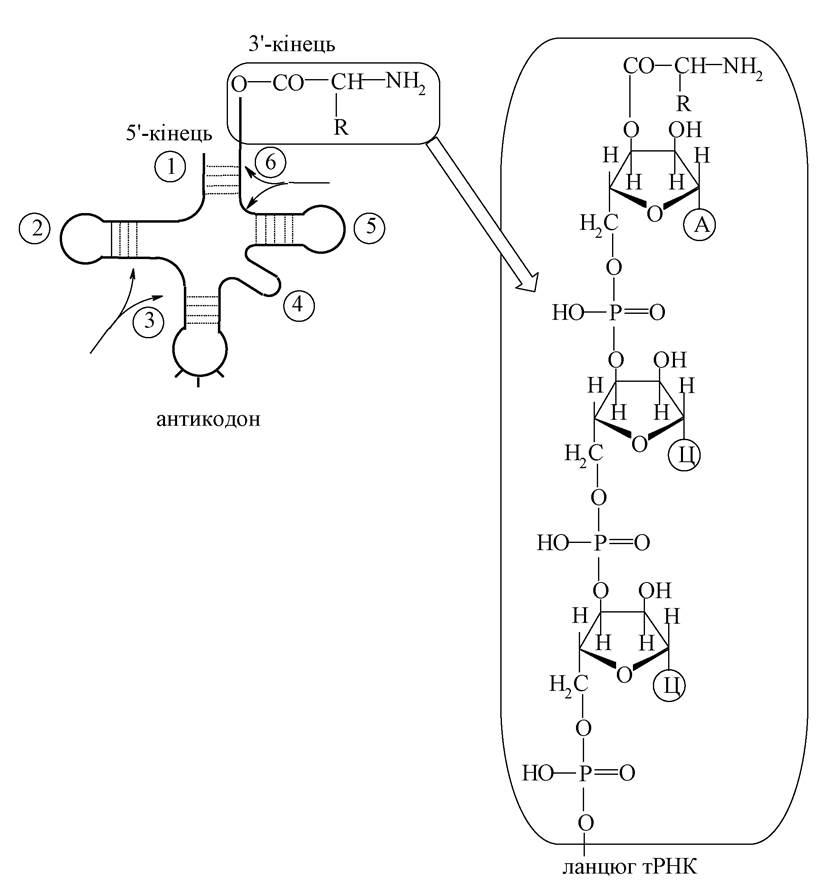
1. 5'-Кінцева ділянка, як правило, у всіх РНК закінчується залишком гуанілової кислоти.
2. Дигідроуридилова гілка містить кілька залишків дигідроуридилової кислоти.
3. Антикодонова гілка містить триплет основ, який називається антикодоном і відповідає певній а-амінокислоті.
4. Додаткова петля розташовується між антикодоновою і універсальною гілками; ця петля має різну довжину для різних тРНК.
5. Універсальна гілка однакова для всіх тРНК і має олігонуклеотидну ділянку ГТЦ.
6. 3'-кінцева ділянка закінчується триплетом УЦА. Це місце зв'язування з а-амінокислотою.
Хімічна сутність перенесення а-амінокислоти за допомогою тРНК полягає в тому, що а-амінокислота ацилює 3'-ОН групу аденілової кислоти, що стоїть на З'-кінці тРНК за наявності АТФ (активатор). Потім тРНК транспортує сполучену з нею а-амінокислоту в рибосому, де за рахунок водневих зв'язків комплементарних основ знаходить кодон у мРНК, що відповідає власному антикодону. мРНК (іРНК) є матрицею для синтезу білка. Вона складається з одного ланцюга, довжина якого визначається довжиною білкової молекули, що синтезується. У полінуклеотидному ланцюгу мРНК рибонуклеотиди утворюють послідовність, яка визначається комплементарними взаємодіями з нуклеїновими основами ДНК, тобто кодону ДНК відповідає антикодон у мРНК. Антикодон у мРНК зазвичай називають кодоном. До мРНК, що знаходиться в рибосомі, прикріплюються своїми антикодоновими ділянками тРНК, що несуть а-амінокислоти. Між наближеними а-амінокислотами утворюється пептидний зв'язок. Рибосомна РНК (рРНК) безпосередньо бере участь у синтезі білків у рибосомах. На частку рРНК доводиться до 80 % від суми клітинних РНК.
Рибосоми - складні структури, що утворюються з чотирьох рРНК і декількох десятків білків. Рибосоми - «фабрики» з виробництва білків. Більш докладно проблему біосинтезу білка буде розглянуто в курсі біологічної хімії.
7. Деякі мононуклеотиди беруть участь не тільки в будові нуклеїнових кислот, але на їх основі в організмі синтезуються інші важливі біологічні речовини. Так, при послідовному фосфорилюванні АМФ утворюються, відповідно АДФ (аденозин-5'-дифосфат) і АТФ (аденозин-5'-трифосфат).
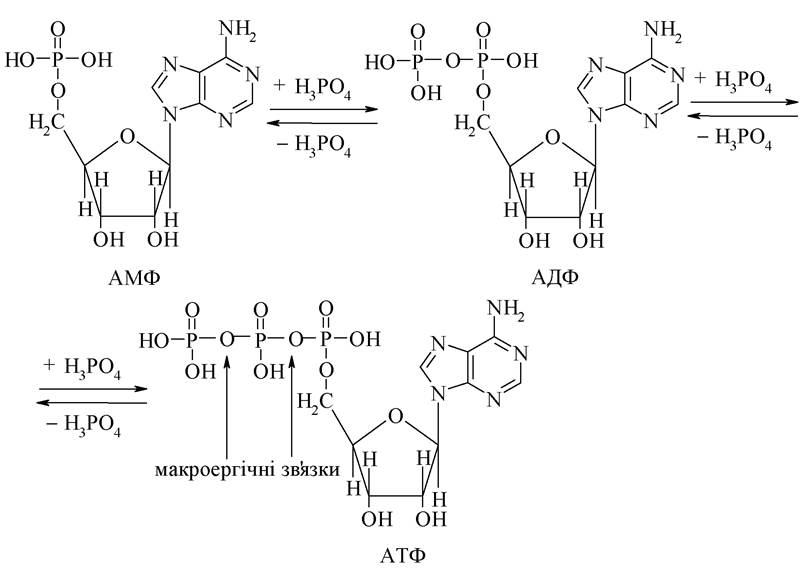
Ці три нуклеотиди здатні зворотньо перетворюватися один в одного. При цьому утворення АДФ і АТФ супроводжується акумулюванням енергії в ангідридних (макроергічних) зв'язках Р-O. При гідролізі АТФ і АДФ ця енергія (32 кДж/моль) виділяється і тому АТФ у біохімічних перетвореннях (спряжених реакціях) виступає як "джерело" енергії. Крім того, АТФ у багатьох біохімічних процесах є активатором жирних кислот, амінокислот, жовчних кислот та ін. сполук шляхом утворення ацилфосфатів.
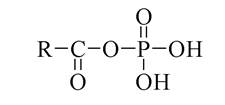
Відомі і інші трифосфати, які виконують аналогічні функції: ЦТФ необхідний для біосинтезу ліпідів, ГТФ - для біосинтезу білків. АМФ бере участь також у будові структур деяких нуклеотидних коферментів. Так, АМФ є складовою частиною коферменту А (КоА-SH).
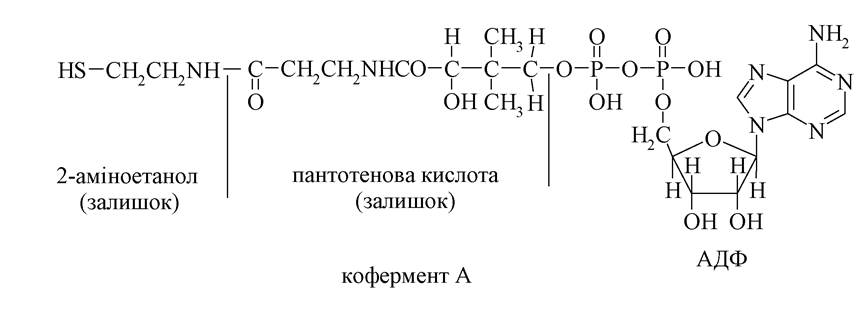
Кофермент А активує карбонові кислоти, перетворюючи їх в реакційноздатні естери тіолів:

8. Іншою важливою групою нуклеотидних коферментів, що містять АМФ, є нікотинамідаденіндинуклетид (НАД+) і нікотинамідаденінди- нуклетидфосфат (НАДФ+). Ці речовини - коферменти окиснювально- відновних ферментів - дегідрогеназ.

Під час біологічного дегідрування (один з можливих випадків окиснення) субстрат втрачає два гідрогена - протон (Н+) і гідрид-іон (Н-). Кофермент НАД+ у ході реакції приєднує гідрид-іон до у-положення піридинового циклу, тому що цей карбон має найбільший позитивний заряд. Унаслідок цього утворюється відновлена форма НАД+ - НАД Н. Цей процес відзначається втратою ароматичності піридинового кільця і перетворенням його в хіноїдну форму, що супроводжується збільшенням енергії системи. У зворотній реакції відбувається окиснення, а енергія, яка при цьому виділяється, витрачається на біохімічні процеси. Наприклад, реакція окиснення молочної кислоти відбувається за участю ферменту лактатдегідрогенази, коферментом якого є НАД+:

Виконайте завдання та перевірте правильність їх розв’язання за еталонами відповідей
Завдання № 1
1. Укажіть структуру урацилу:
2. Пуринові азотисті основи утворюють N-глікозидний зв'язок з моносахаридом за рахунок до атома нітрогену в положенні:
А.3. В 1. С. 7. D. 9.
3. НАД+ - кофермент ферментів:
А.Декарбоксилази. В. Дегідрогенази. С. Трансферази. D. Каталази.
Еталони відповідей: 1-А; 2-D; 3-В.
Завдання № 2
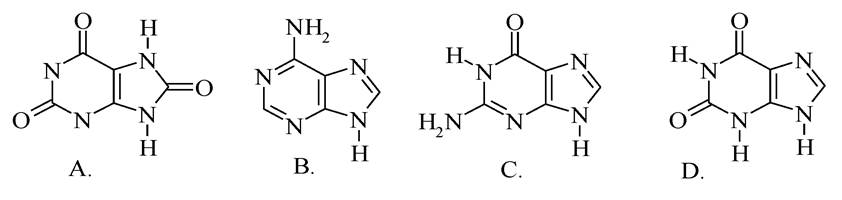
1.Укажіть формулу аденіну:
2. Мононуклеотид - продукт фосфорилювання нуклеозиду відповідно до -ОН групи моносахариду в положенні:
А.2'. В 3’. С. 5'. D. 3'і 5'.
3. Кофермент А в організмі необхідний:
A. Для активації залишків карбонових кислот.
B. Для участі в процесі декарбоксилювання.
C. Для участі в процесі карбоксилювання.
D. Для участі в процесі синтезу білків.
Еталони відповідей: 1-В; 2-С; 3-А.
Завдання № 3
1. Які продукти утворюються в результаті повного гідролізу тимідилової кислоти?
A. Тимідин і фосфорна кислота.
B. Тимін, рибоза і фосфорна кислота.
C. Тимін, дезоксирибоза і фосфорна кислота.
D. Тимін і дезоксирибоза.
2. Яка кількість пуринових і піримідинових основ у ДНК?
A. Однакова.
B. Кількість піримідинових основ більша, ніж пуринових.
C. Кількість піримідинових основ менша, ніж пуринових.
D. Їх співвідношення не має значення.
3. Макроергічною сполукою в організмі є:
А. НАД+. В. НАДФ+. С. Кофермент А. D. АТФ.
Еталони відповідей: 1-С; 2-А; 3-D.