Основы биохимии - А. А. Анисимов 1986
Нуклеиновые кислоты
Общая характеристика строения нуклеиновых кислот
4.2.1. Компоненты нуклеиновых кислот. При полном гидролизе нуклеиновых кислот образуются пуриновые и пиримидиновые азотистые основания, моносахарид пентоза (рибоза или дезоксирибоза) и фосфорная кислота. В выделении и доказательстве присутствия рибозы в нуклеиновых кислотах большую роль сыграли работы советского физиолога и биохимика Е. С. Лондона (1929).
Все нуклеиновые кислоты делятся на два типа в зависимости от того, какой моносахарид входит в их состав. Нуклеиновая кислота называется рибонуклеиновой (РНК), если в ее состав входит рибоза, или дезоксирибонуклеиновой (ДНК), если в ее состав входит дезоксирибоза.
Пентозы в нуклеиновых кислотах присутствуют всегда в ß-D-фуранозной форме:

Углеродные атомы пентозы нумеруются цифрами со знаком «штрих» для того, чтобы можно было отличить их от атомов азотистого основания (например, 5-й углеродный атом обозначают С-5' или 5'). Небольшое отличие — присутствие Н— вместо ОН— у С-2' рибозы является одной из основных причин существенных различий в свойствах ДНК и РНК. Предполагают, что замена ОН— на Н— упрочняет связь между 2-м и 3-м углеродом рибозы. Эго в целом увеличивает прочность молекулы ДНК, ее консервативность как хранителя наследственности. Отсутствие кислорода у С-2' дезоксирибозы способствует компактности укладки молекулы ДНК, что важно для вмещения ее гигантских молекул в малом объеме клеток и ядер.
Пуриновые и пиримидиновые азотистые основания, входящие в состав нуклеиновых кислот, являются производными ароматических гетероциклических соединений — пурина и пиримидина. Молекула пурина состоит из двух конденсированных колец: пиримидина и имидазола. Среди пуриновых азотистых оснований главную роль играют аденин (А) и гуанин (Г), а среди пиримидиновых оснований — цитозин (Ц), урацил (У), тимин (Т; 5-метилурацил);

В состав ДНК входят аденин, гуанин, цитозин, тимин; в РНК вместо тимина присутствует урацил. Ниже приведены одинаковые и отличающиеся компоненты ДНК и РНК.
Одинакорые компоненты |
Отличающиеся компоненты |
|
Аденин |
ДНК |
РНК |
Гуанин |
Дезоксирибоза |
Рибоза |
Цитозин |
Тимин |
Урацил |
Кроме главных азотистых оснований в нуклеиновых кислотах присутствуют в небольших количествах необычные — минорные основания. Так, в состав ДНК высших организмов входит 5-метил-цитозин, содержание которого у высших растений намного превышает его содержание у животных. В ДНК ряда бактерий встречаются небольшие количества 6-метиладенина и 5-метилцитозина. Эти метилированные основания защищают «свои» ДНК от расщепления ферментами — ДНКазами. В ДНК Т-четных фагов Е. coli цитозин заменен на 5-гидроксиметилцитозин. В некоторых случаях его гидроксиметильная группа соединена с глюкозой. Особенно много минорных компонентов содержится в транспортных РНК: тиоурацил, дигидроурацил, псевдоуридин1, ксантин (2,6-диоксипурин), гипоксантин (6-оксипурин), ацетилцитозин, оротовая кислота и другие, всего около 60.
1 Псевдоуридин — это минорный нуклеозид, в котором рибоза присоединяется не к 1-му, а к 5-му атому урадила.

Трехмерная структура различных пуриновых и пиримидиновых оснований была исследована методом рентгеноструктурного анализа. Молекулы пиримидинов имеют плоское строение, а молекулы пуринов — почти плоское. Все они, кроме аденина, существуют в таутомерных формах. Так, урацил может находиться в форме или лактима, или лактама:
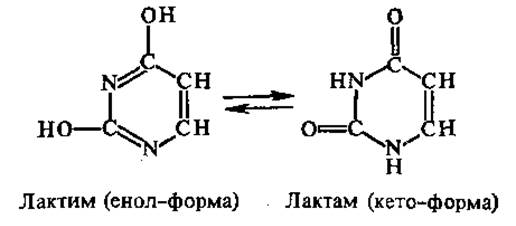
В составе нуклеиновых кислот все оксопроизводные азотистые основания присутствуют в форме лактамов. Азотистые основания поглощают свет в ультрафиолетовой области спектра с длинами волн 200—300 нм и максимумом около 260 нм. Это свойство используют при количественном определении нуклеиновых кислот.
В процессе обмена веществ растений и животных пуриновые основания образуют ряд продуктов: мочевую кислоту, кофеин, теобромин. В последние годы некоторые синтетические пиримидины широко используют в качестве биологически активных соединений.
4.2.2. Нуклеозиды и нуклеотиды. В нуклеиновых кислотах пуриновые авотистые основания через 9-й атом, а пиримидиновые — через 1-й образуют N-гликозидную связь с пентозой рибозой или 2'-дезоксирибозой.
Такие соединения, в которых азотистые основания связаны с рибозой или дезоксирибозой, называются нуклеозидами, а их фосфорные эфиры — нуклеотидами. Если аденин присоединяется к рибозе, то получается нуклеозид аденозин. Если к аденозину присоединяется остаток фосфорной кислоты в 5'-положении, то образуется 5'-адениловая кислота, или аденозин-5'-монофосфат, если в 3'-положении — то 3'-адениловая кислота, или аденозин-3'-монофосфат. Номенклатура нуклеозидов и нуклеотидов приведена в табл. 4.1. Фосфат может этерифицировать сахар также в 2'- или 3'-положении.
Все нуклеотиды — сильные кислоты, так как остаток фосфорной кислоты легко диссоциирует. К нуклеозидмонофосфату могут присоединиться посредством фосфоангидридной связи еще один или два остатка фосфорной кислоты. При этом образуются нуклеозидди- и нуклеозидтрифосфаты. Если в состав нуклеозида входит дезоксирибоза, то перед названием соответствующего нуклеотида ставится приставка дезокси, например, д-АТФ-дезоксиаденозин-5'-трифосфат.
4.2.3. Строение полинуклеотидной цепи Нуклеотиды — это повторяющиеся мономерные единицы олигонуклеотидов и полинуклеотидов. Олигонуклеотиды построены из нескольких мономеров, полинуклеотиды — из многих. Нуклеиновые кислоты представляют собой полинуклеотиды, построенные из мономеров — нуклеотидов, число которых колеблется от 7 десятков до сотен миллионов.
Молекулы нуклеиновых кислот всех типов живых организмов — линейные полимеры, не имеющие разветвлений, что доказано с помощью ряда методов (химических, ферментативных, электронной микроскопии). Наличие так называемых палиндромных шпилек (см. разд. 4.3.1), образующих своеобразные боковые отростки от линейной цепи ДНК, не следует считать нарушением ее линейности, так как в пределах каждой шпильки линейность структуры сохраняется. При суперспирализации ДНК также могут возникать своеобразные структуры, однако это тоже будут различные изгибы непрерывно продолжающейся линейной цепи. Роль мостика между нуклеотидами выполняет 3', 5'-фосфодиэфирная связь, соединяющая С-3' D-рибозы (или 3'-дезоксирибозы) одного нуклеотида и С-5' другого (рис. 4.1).
Таблица 4.1. Номенклатура нуклеозидов и нуклеотидов
Нуклеотиды |
|||
Азотистые основания |
Нуклеозиды |
полное название |
сокращенное название |
Аденин |
Аденозин |
Адениловая кислота, аденозинмоно-фосфат |
АМФ |
Гуанин |
Гуанозин |
Гуаниловая кислота, гуанозинмоно-фосфат |
ГМФ |
Цитозин |
Цитидин |
Цитидиловая кислота, цитидинмоно-фосфат |
ЦМФ |
Урацил |
Уридин |
Уридиловая кислота, уридинмоно-фосфат |
УМФ |
Тимин |
Тимидин |
Тимидиловая кислота, тимидинмоно-фосфат |
ТМФ |
В связи с этим полинуклеотидная цепь имеет определенное направление. На одном ее конце остается свободной 5'-ОН-группа (начало цепи), на другом — 3'-ОН-группа (конец цепи). Концевые гидроксильные группы могут быть этерифицированы фосфатом. Полное написание полинуклеотидных цепей сложно и громоздко, поэтому применяют схематическое изображение. Вертикальными прямыми линиями обозначают углеродные цепи сахаров с основаниями, присоединенными к С-1' диагоналями с буквой Р в середине — фосфодиэфирные связи между атомом С-3' (вблизи середины прямой линии) и С-5' (конец следующей линии). Используют также буквенную систему: буквами А, Г, Ц, Т, У обозначают азотистые основания, буквой ф — фосфатную группу. Если буква ф находится слева, этерифицирована группа при С-5', справа — этерифицирована группа при С-З' (рис.4.1). Например, фА — аденозин-5'-фосфат, Аф — аденозин-3'-фосфат.
1 Иногда употребляется также и термин нить.

Рис. 4.1. Строение полинуклеотидной цепи РНК:
А, Г, Ц — азотистые основания; І — полная, ІІ — сокращенная форма записи
Полинуклеотидная цепь несет множество фосфатных групп, которые легко диссоциируют, вследствие чего она приобретает отрицательный заряд. В связи с этим нуклеиновые кислоты в клетке во многих случаях связываются с основными белками, образуя нуклеопротеины. РНК входят в состав рибонуклеопротеинов
(РНП), ДНК — дезоrсирибонуклеопротеинов (ДНИ).
Нуклеопротеины экстрагируют обычно из клеток 1 М раствором нейтральных солей. Существенным моментом выделения нуклеиновых кислот является отщепление от нуклеопротеинов белков и их удаление. Этого можно достичь использованием в качестве растворителя белков фенола или смеси хлороформа с изоамиловым спиртом. С этой целью применяют также детергенты (додецилсульфат натрия) или расщепление белков протеиназами. Из последних особенно эффективна проназа (протеиназа Streptomyces griseus). После отделения белка нуклеиновую кислоту можно осадить, медленно прибавляя этанол. При препаративном выделении нуклеиновых кислот их получают в виде бариевых, натриевых или литиевых солей, в которых отрицательные заряды фосфатных групп экранированы катионами.
В препаратах ДНК обнаруживаются в малых количествах Si, Mg, Ca, Sr и ряд других микроэлементов, которые, по-видимому, участвуют в стабилизации структуры. Есть предположение, что кремний в форме кремниевой кислоты может в некоторых случаях заменять фосфатные остатки в молекуле ДНК.