Основы биохимии - А. А. Анисимов 1986
Витамины
Жирорастворимые витамины
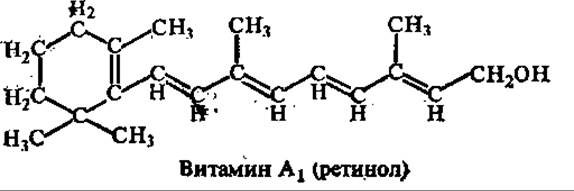
Витамин А (антиксерофтальмический, ретинол, аксерофтол). Известны три витамина этой группы: А1, А2 (у них все двойные связи находятся в транс-конфигурации) и неовитамин А (цис-форма витамина А1). Витамин А2 отличается от A1 присутствием дополнительной двойной связи в кольцевой части молекулы.
Ранним симптомом авитаминоза А является ослабление темновой адаптации, вплоть до полной утраты зрения в сумерках — куриная слепота. Одно из основных проявлений авитаминоза А заключается в системном поражении эпителиальной, ткани. На этой основе, в частности, происходит поражение роговой оболочки глаза. Нарушается строение выстилающего ее защитного эпителия, он подвергается ороговению, высыхает, теряет прозрачность — развивается ксерофтальмия (от греч. ксерос — сухой, офтальмос — глаз), за которой следует кератомаляция (размягчение роговицы) с последующим некрозом и изъязвлением. После заживления этих повреждений остается рубец — бельмо, преграждающий доступ света к сетчатке. Повышенному ороговению подвергается и эпителий кожи, что способствует возникновению кожных болезней. Особенно опасны изменения эпителия, выстилающего слизистые оболочки дыхательных путей (бронхиты), кишечника (колиты).
При А-витаминной недостаточности ослабевают механизмы иммунитета. Избыточное введение витамина А может вызвать гипервитаминоз, интоксикацию. Особенно тяжело она протекает у детей. Основными источниками витамина А служат яйца, сливки, сметана, коровье молоко, сливочное масло, почки, печень крупного рогатого скота и печень трески. Рекомендуемое суточное потребление витамина А для взрослого человека — 1 мг. Витамин А в высших растениях и микроорганизмах не синтезируется, но у них образуются его предшественники — каротиноиды. Их особенно много в моркови, помидорах, шпинате, перце. Биосинтез каротинов, как и всех терпенов, осуществляется из ацетил-КоА через мевалоновую кислоту (см. разд. 8.8.3).
Структурные изомеры каротина (a-, ß-, у-каротины) способны превращаться в организме человека и животных в витамин А. В процессе превращения каротина в витамин А (ретинол) окислительное расщепление углеродной цепи провитамина происходит двумя путями: по периферическим двойным связям и по центральной двойной связи. В организме ретинол окисляется в ретиналь и ретиноевую кислоту при участии соответствующих дегидрогеназ и НАД. Ретиналь занимает ключевое положение в обмене витамина А и легко подвергается энзиматическому обратимому восстановлению в ретинол и необратимому окислению в ретиноевую кислоту.
Наиболее специфической функцией витамина А является его участие в процессах фоторецепции. Фоточувствительный пигмент, находящийся в наружном сегменте палочек сетчатки (ответственны за сумеречное зрение) человека, наземных позвоночных и морских рыб, — родопсин — является хромопротеином, состоящим из хромофорной группы — витамина А — альдегида (ретиналь) и белка опсина. Связь между ними осуществляется через альдегидную группу ретиналя и свободную NН2-группу белка с образованием шиффова основания. Под действием света 11-цис-ретиналь отщепляется от родопсина и одновременно переходит в транс-форму. Обесцвеченная в результате отщепления ретиналя, молекула родопсина запускает сложную цепь ферментных реакций в зрительной клетке — ферментативный каскад усиления слабого светового сигнала. Обесцвечивание родопсина связано также с повышением проницаемости фоторецепторной мембраны. В результате всех этих процессов происходит возбуждение зрительного нерва.
Есть сведения, что важную роль в зрительной рецепции играют цАМФ и аденилатциклаза. Установлена активация светом адени- латциклазы у некоторых животных. Дофамин, являющийся одним из главных нейромедиаторов в сетчатке, активирует аденилатциклазу, повышает уровень содержания цАМФ в интактной ткани. цАМФ зависимое фосфорилирование родопсина, по мнению некоторых авторов, может быть причиной гиперполяризации мембраны и возникновения первичного нервного импульса. Затем транс-ретиналь через транс-ретинол вновь изомеризуется в цис-ретиналь и соединяется с опсином.
Поглотившие квант света цис- или транс-изомеры переходят из основного (невозбужденного) состояния в синглетное (электронновозбужденное состояние). В этом состоянии между атомами углерода имеется простая (не двойная) связь, что крайне облегчает изомеризацию — цис-транс-переходы. Часть ретиналя при этих превращениях разрушается, для его образования требуются новые молекулы витамина А. Вследствие недостатка последнего теряется способность видеть вечером и ночью.
В ходе биологической эволюции 11-цис-ретиналь сохранился в качестве хромофорной группы зрительных пигментов всех позвоночных животных и беспозвоночных — моллюсков, ракообразных, насекомых. Даже у эволюционно древних галобактерий, находящихся, видимо, на слепой ветви эволюции, в качестве светочувствительной рецепторной молекулы функционирует ретин альсодержащий мембранный белок.
Важное проявление механизма действия витамина А заключается в его участии в регуляции проницаемости мембран, а также в транспорте моносахаридов, необходимых для биосинтеза гликопротеинов. Витамин А оказывает несомненное влияние на усвоение белка пищи и его обмен в организме, а также на некоторые стороны обмена липидов, в том числе убихинона, сквалена, холестерина и частично фосфолипидов.
Предполагают участие витамина А в окислительно-восстановительных реакциях: поскольку в молекуле витамина А имеются двойные связи, он способен образовывать пероксиды, повышающие скорость окисления других соединений.
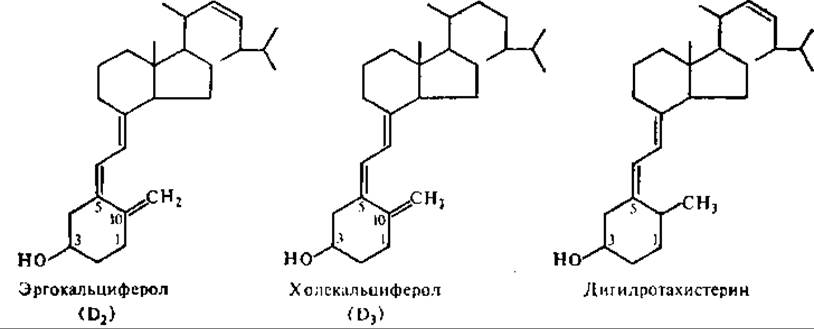
Витамин D (антиоахитический. кальциферолы). Название кальциферолы объединяет группу родственных соединений, обладающих антирахитической активностью. Важнейшие среди них — холекальциферол (витамин D3), эргокальциферол (витамин О2), дигидроэргокальциферол (витамин D4).
Основное отличие структуры кальциферолов от других стероидных соединений состоит в размыкании одного кольца фенантрена, появлении в молекуле трех сопряженных двойных связей и метиленовой группы вместо метальной.
Недостаток витамина D приводит к возникновению рахита. При этом заболевании задерживается зарастание швов между костями черепа, которые избыточно разрастаются; увеличиваются лобные бугры. Деформируются и другие кости. Вследствие недостаточного окостенения реберных хрящей грудная клетка приобретает неправильную форму. Кости ног под влиянием тяжести тела искривляются. Мышцы становятся дряблыми, их тонус понижается, увеличивается живот. Рахит тормозит общее развитие ребенка. Задерживается прорезывание зубов, а сами зубы легко разрушаются, часты желудочно-кишечные расстройства, развивается малокровие, ребенок становится легко подвержен другим заболеваниям.
Большие дозы витамина могут вызвать гипервитаминоз D, который проявляется в резком похудании, остановке роста, подъеме кровяного давления, повышении температуры, резких болях в суставах, судорогах, затруднении дыхания. Источниками витамина D в пище являются рыбий жир, печень трески, икра, яичные желтки, сливочное масло. Суточное потребление витамина D детьми до 6 лет должно составлять от 500 до 1000 ME, а в более старшем возрасте во взрослом состоянии — 100 ME. Однако в поддержании необходимого общего содержания витамина D в организме важнейшую роль играет его эндогенное образование из 7-дегидрохолестерина под влиянием ультрафиолетовой части света.
Провитамином холекальциферола является образующийся из холестерина 7-дигидрохолестерин, провитамином эргокальциферола — эргостерин, присутствующий в растениях и микроорганизмах (особенно много в дрожжах). Противорахитным действием обладает и дигидротахистерин — каталитически восстановленный тахистерин, образующийся при ультрафиолетовом облучении эргостерина.
В зеленых растениях витамин D не синтезируется, но они являются поставщиками 7-дегидрохолестерина, необходимого для образования витамина D. Витамин D выполняет свои специфические функции в обмене веществ не в виде холе- или эргокальциферола, а в форме образующихся из них активных метаболитов, важнейшим из которых является 1,25-диоксихолекальциферол. Холекальциферол в печени превращается под действием митохондриальных ферментов при участии НАДН и О2 в 25-оксихолекальциферол. Последний гидроксилируется в почках с образованием 1,25-диоксихолекальциферола. Основные функции витамина D в организме связаны с обеспечением транспорта Са и Р через биологические мембраны. Участие витамина D в следующих процессах можно считать хорошо установленным: 1. Перенос ионов Са и Р через эпителиальные клетки слизистой тонкого кишечника в процессе их всасывания. Витамин D вступает в комплекс с Са-связывающим белком в кишечнике. Существует гипотеза, согласно которой один из белков слизистой кишечника под влиянием 1,25-диоксихолекальциферола приобретает способность активно связывать Са. По другому предположению, 1,25-диоксихолекальциферол влияет на биосинтез мРНК для трансляции Са-связывающего белка. 2. Мобилизация кальция из скелета путем рассасывания предобразованной костной ткани. 3. Реабсорбция фосфата и кальция в почечных канальцах. В итоге витамин D обусловливает оптимальное содержание Са и Р в плазме крови, необходимое для минерализации костной ткани.
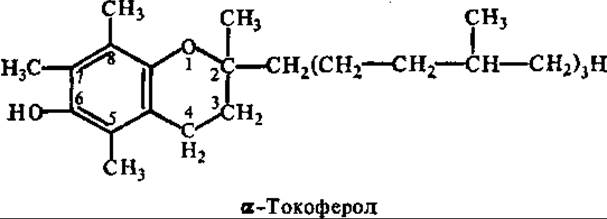
Витамин Е (антистерильный, токоферолы). К группе витамина Е относятся метальные производные токола и токотриенола. Индивидуальные токоферолы, обозначаемые греческими буквами а, ß, у и δ, отличаются друг от друга количеством и положением дополнительных метальных заместителей в ароматическом кольце 6-оксихромана. a-, ß-, у- и δ-Токотриенолы — аналоги соответствующих токоферолов и отличаются от них структурой боковой полиизопреноидной цепи. Последняя у всех жирорастворимых витаминов формируется из ацетил-КоА-через мевалонат и изопентенилпирофосфат.
Для недостаточности витамина Е характерно следующее: 1. Резорбция плодов при беременности. 2. Дегенерация семенников у самцов: снижение подвижности сперматозоидов и прогрессирующая дегенерация зародышевого эпителия с атрофией и уменьшением массы семенников. 3. Мышечная дистрофия с коагулирующим или гиалиновым некрозом мышечных клеток, атаксией и параличами. Это одно из основных проявлений недостаточности витамина Е у кроликов, морских свинок и многих видов домашних животных. 4. Макроцитарная (крупноклеточная) анемия у обезьян и человека со снижением продолжительности жизни эритроцитов и нарушением эритропоэза в костном мозгу. 5. Повышенная чувствительность эритроцитов к перекисному гемолизу in vitro — одно из наиболее универсальных проявлений недостаточности витамина Е, характерное, видимо, для всех видов животных.
Глубина и характер проявлений недостаточности витамина Е существенно зависят от других компонентов рациона. Дефицит белка, Se, избыточное содержание жиров, особенно ненасыщенных, добавки солей Fe, Ag и некоторых других металлов могут значительно ускорять и углублять развитие недостаточности витамина Е. Такие признаки авитаминоза, как некроз печени у крыс, миопатии у цыплят, возникают только в том случае, если дефицит витамина Е сочетается с недостаточным содержанием в рационе Se. Токоферолы широко распространены в природе, особенно в растениях и растительных продуктах. Наиболее богаты ими растительные масла (из пшеничных зародышей, кукурузное, хлопкое, подсолнечное). Суточная норма потребления здоровыми взрослыми людьми витамина Е составляет 15 ME.
Конкретный механизм действия витамина Е на молекулярном уровне окончательно не расшифрован. Одна из наиболее тщательно разработанных гипотез, позволяющих на единой основе объяснить многочисленные и разнообразные проявления недостаточности витамина Е, — антиоксидантная гипотеза. В соответствии с ней токоферолы выполняют в живых тканях роль биологических антиоксидантов, инактивирующих свободные радикалы и тем самым препятствующих развитию нерегулируемых, неферментативных, цепных свободнорадикальных процессов перекисного окисления ненасыщенных тканевых липидов молекулярным кислородом. Поскольку ненасыщенные липиды являются компонентами липопротеинов мембран клеток и субклеточных органелл, то усиление их перекисного окисления при снижении концентрации токоферола в тканях приводит к повреждению структуры, нарушению проницаемости и функциональной активности клеточных и субклеточных мембран. Этот дефект и лежит в основе многообразных биохимических, морфологических и клинических проявлений недостаточности витамина Е.
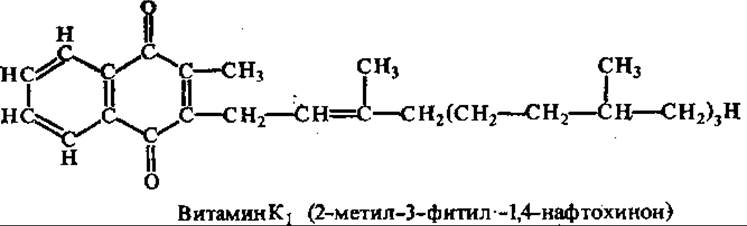
Витамин К (антигеморрагический, филлохиноны, менахиноны). Витамины группы К широко распространены в природе и представлены двумя рядами хинонов — филлохинонами (витаминами К1-ряда) и менахинонами (витаминами К1-ряда). Основой молекулы тех и других является 1,4-нафтохинон.
Филлохиноны отличаются от менахинонов структурой боковой цепи, у менахинонов она полиизопреноидная, число изопреновых остатков указывается в названии (например, менахинон-4). Филлохиноны и их деметилированные производные обнаружены в растениях, менахиноны синтезируются различными бактериями или являются продуктами трансформации нафтохинонов в организме. В животных тканях нафтохиноны представлены филлохинонами и менахинонами алиментарного происхождения (поступают с пищей), а также менахинонами, которые образуются в организме из филлохинона. Кроме того, менахинон-4 может образовываться в организме при введении синтетических нафтохинонов, например викасола, синкавита, менадиона и др.
При авитаминозе К появляются подкожные и внутримышечные кровоизлияния (геморрагии), снижается скорость свертывания крови. Первичная недостаточность витамина К у взрослых людей наблюдается редко. Объясняется это тем, что потребность в нем обычно обеспечивается поступлением с пищевыми продуктами, где он широко распространен, и за счет его синтеза кишечными бактериями. Дефицит витамина К нередко наблюдается у новорожденных детей из-за низкого его содержания в молоке и отсутствия в кишечнике синтезирующей его микрофлоры.
Вторичная недостаточность витамина К возникает вследствие болезней печени, особенно обтурационной желтухи, хронических заболеваний кишечника, при лечении сульфаниламидами и антибиотиками, угнетающими кишечную микрофлору, а также под влиянием лечения препаратами, являющимися антагонистами витамина К. Богатые источники витамина К — зеленые растения, где он содержится в хлоропластах в виде филлохинона. Синтез витамина К в растениях связан с их фотосинтетической функцией. Особенно богатым шпинат, капуста, тыква. В микроорганизмах и животных тканях присутствуют различные формы витамина К2. В отличие от микроорганизмов и растений, обладающих способностью синтезировать витамин К, для человека и высших животных он является экзогенным фактором. Остро нуждаются в нем птицы.
У человека потребность новорожденных в витаминах составляет от 2 до 12 мкг/сут. Всасывание витаминов К происходит в тонком кишечнике. Природным витаминам К1 = К2-ряда, обладающим липофильными свойствами, требуется присутствие желчных кислот и панкреатической липазы, тогда как водорастворимые формы витамина К в этом не нуждаются. Биологически активной формой витамина К в организме человека и животных является менахинон-4, так как все другие формы витамина К трансформируются в нее. Конкретный молекулярный механизм действия нафтохинонов окончательно не выяснен. Наиболее вероятными считают следующие гипотезы.
1. Участие витамина К в окислительном фосфорилировании. Из митохондрий микробных и растительных клеток, а также из хлоропластов были выделены филлохинонредуктаза, менадионредуктаза, в которых витамин К выполняет коэнзимную функцию. Нафтохиноны наряду с бензохинонами присутствуют в составе фотосинтетической системы и участвуют в переносе световой энергии к хлорофиллу. Признано также участие витамина К в окислительном фосфорилировании у некоторых бактерий. Однако данных, доказывающих специфическое участие витамина К в процессах окислительного фосфорилирования в животном организме, пока нет.
2. Действие витамина К на генетическом уровне. На основании опытов с использованием актиномицина D, блокирующего биосинтез белков на стадии образования мРНК, был сделан вывод, что витамин К вовлекается в биосинтез факторов свертывания крови, стимулируя ДНК-зависимый синтез соответствующей мРНК, и что антагонизм между витамином К и дикумарином осуществлялется на уровне гена-регулятора.
3. Участие витамина К в синтезе прокоагулянтов (факторов свертывания крови II, VII, IX и X). Витамины К участвуют в посттрансляционном карбоксилировании остатков глутаминовой кислоты в составе перечисленных белков (факторов свертывания), что необходимо для их активного функционирования. Карбоксилирование остатков глутаминовой кислоты осуществляется микросомальной карбоксилазой. Солюбилизированная карбоксилаза нуждается в НАДН, витамине К, НСО3 и О2. Витамин К обладает коферментной функцией, которая реализуется в форме 2,3-эпоксида. Окисление в 2,3-эпоксид происходит под действием специфической оксидоредуктазы, восстановление — под действием НАДФН-диафоразы. Ни одна из приведенных гипотез в настоящий момент не может считаться всеобъемлющей и единственно верной.