Биохимия и молекулярная биология - Белясова Н.А. 2002
Метаболизм. Процессы, приводящие к запасанию энергии
Цикл трикарбоновых кислот
Химизм ЦТК
Реакции цикла трикарбоновых кислот осуществляются у эукариот в митохондриях — там же, где функционируют системы, поставляющие «топливо» для ЦТК: ß-окисление жирных кислот, окислительное декарбоксилирование пирувата. Считается, что ферменты, принимающие участие в ЦТК, сгруппированы в особый каталитический комплекс, расположенный между внутренними мембранами митохондрий. Структура комплекса такова, что промежуточные продукты цикла последовательно переходят от одного фермента к другому, не высвобождаясь в матрикс. В этом случае скорость процесса значительно увеличивается.
Все реакции ЦТК можно условно разделить на 4 стадии: 1) акцептирование ацетил-СоА и образование изоцитрата; 2) окислительное декарбоксилирование изоцитрата; 3) окислительное декарбоксилирование a-кетоглутарата; 4) регенерация акцептора ацетил-СоА.
Акцептором ацетил-СоА является оксалоацетат (щавелевоуксусная кислота, ЩУК). Это четырехуглеродное соединение конденсируется с двухуглеродным компонентом ацетил-СоА при участии фермента цитратсинтетазы. Данная реакция альдольной конденсации протекает ступенчато, с образованием промежуточного продукта — цитрил-СоА, гидролиз которого сдвигает направление реакции в сторону образования цитрата (рис. 11.2).
Чтобы могло осуществиться окислительное декарбоксилирование шестиуглеродного соединения, цитрат (лимонная кислота) должен изомеризоваться в изоцитрат. Этот процесс катализируется аконитазой и представляет собой реакцию дегидратации с последующей гидратацией.
Окислительное декарбоксилирование изоцитрата катализируется изоцитратдегидрогеназой, которая использует NAD в качестве кофермента. Изоцитрат окисляется в ß -кетокислоту, оксалосукцинат, который быстро теряет карбоксил, будучи связанным с ферментом, и превращается в a-кетоглутарат (рис. 11.3).
На третьей стадии (рис. 11.4) a-кетоглутарат также подвергается окислительному декарбоксилированию. Эту реакцию катализирует организованный ансамбль ферментов — a-кетоглутарат-дегидрогеназный комплекс. Состав ансамбля сходен с комплексом, катализирующим окислительное декарбоксилирование пирувата, и оба процесса имеют множество общих деталей. Продуктом реакции является четырехуглеродное активированное соединение — сукцинил-СоА. Макроэргическая связь сукцинильного тиоэфира расщепляется с участием сукцинаттиокиназы, и это сопряжено с фосфорилированием GDP (у млекопитающих) или АТР (у бактерий, высших растений). Образование высокоэнергетической связи GTP или АТР при гидролизе сукцинил-СоА является единственным случаем субстратного фосфорилирования, который имеет место в ЦТК.
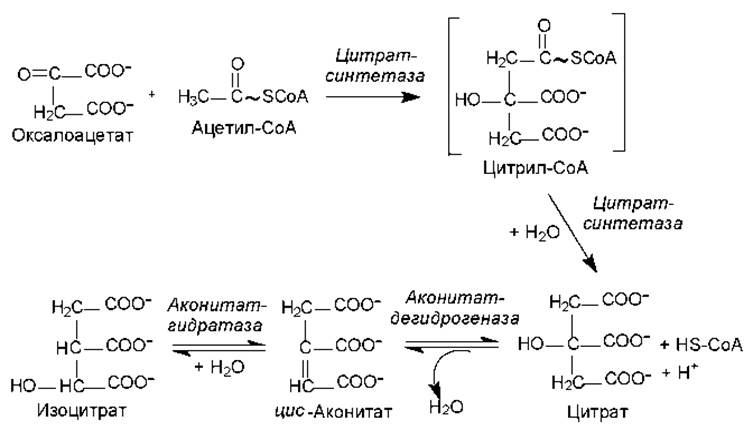
Рис. 11.2. Первая стадия ЦТК: образование изоцитрата
Образовавшийся на предыдущей стадии сукцинат (янтарная кислота) в ходе нескольких реакций (окисление, гидратация, окисление) превращается в оксалоацеат (рис. 11.5). Данная последовательность превращений представляет собой процесс ß-окисления, который уже был рассмотрен по отношению к жирным кислотам в главе 9. Реакции последней стадии ЦТК представляют собой регенерирование оксалоацетата. Окисление сукцината катализирует сукцинатдегидрогеназа. Этот фермент непосредственно связан с цепью переноса электронов, и в отличие от других ферментов ЦТК является интегральным белком внутренней мембраны митохондрий.
Продукт окисления сукцината — фумарат — гидратируется фумаразой. Происходит стереоспецифическое присоединение водорода и гидроксила с образованием L-малата (яблочной кислоты). Наконец, L-малат окисляется в оксалоацетат малатдегидрогеназой.
Полностью циклическая система реакций ЦТК представлена на рис. 11.6. Многие промежуточные вещества ЦТК служат предшественниками в биосинтетических реакциях. Например, оксалоацетат и a-кетоглутарат используются в биосинтезе аминокислот. Оксалоацетат включается в процесс глюконеогенеза (образование глюкозы из неуглеводных субстратов). Большинство углеродных атомов в порфиринах ведут свое происхождение от сукцинил-СоА. Однако потребление промежуточных продуктов ЦТК для целей биосинтеза обязательно должно сопровождаться их восполнением, в противном случае цикл прервется. В норме между реакциями, за счет которых определенные промежуточные продукты удаляются из цикла и, наоборот, поступают в цикл, существует состояние динамического равновесия.

Рис. 11.3. Вторая стадия ЦТК: окислительное декарбоксилирование изоцитрата
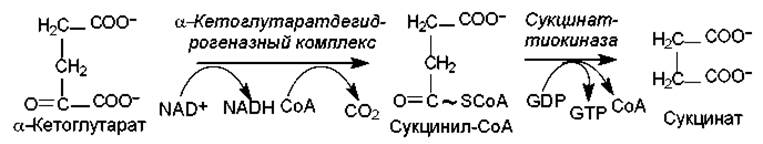
Рис. 11.4. Третья стадия ЦТК: окислительное декарбоксилирование a-кетоглутарата
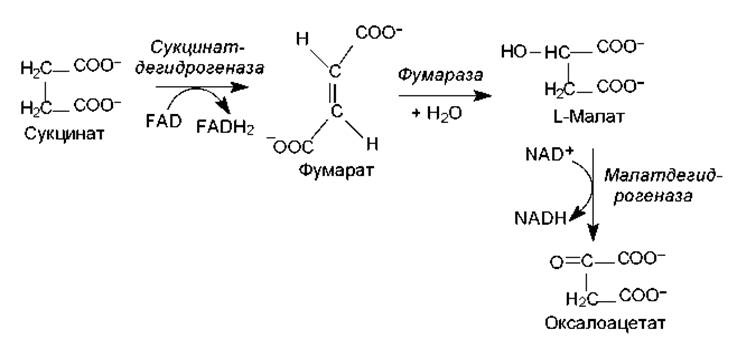
Рис. 11.5. Заключительная стадия ЦТК: регенерирование оксалоацетата
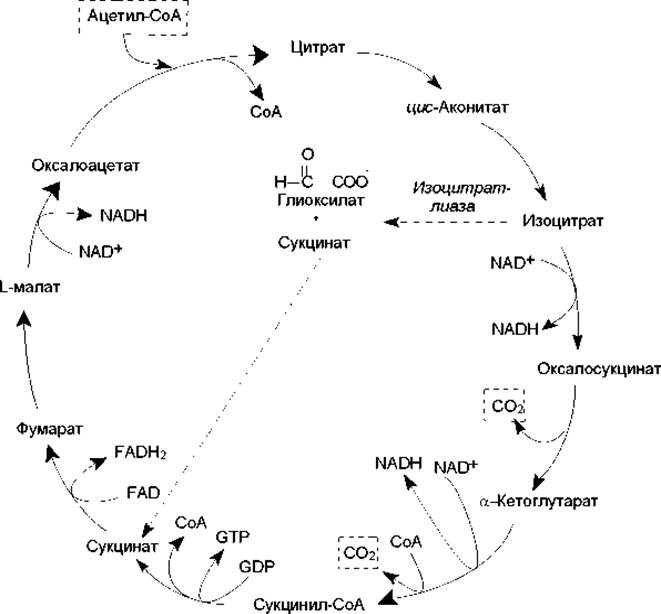
Рис. 11.6. Цикл трикарбоновых кислот. Пунктирными линиями обозначены превращения глиоксилатного цикла
Восполняющие запас промежуточных продуктов ЦТК реакции носят название анаплеротических. Одной из основных анаплеротических реакций клеток млекопитающих является карбоксилирование пирувата с образованием оксалоацетата. Катализирует это превращение аллостерический фермент — пируваткарбоксилаза. Его активатором служит субстрат ЦТК — ацетил-СоА. В условиях накопления ацетил-СоА стимулируется активное карбоксилирование пирувата и увеличивается количество оксалоацетата, который служит непосредственным акцептором ацетил-СоА.
В клетках бактерий и растений восполнение оксалоацетата осуществляет другой фермент — фосфоенолпируват-карбоксилаза.