Биологическая химия - Березов Т. Т., Коровкин Б. Ф. 1998
Обмен сложных белков
Обмен нуклеиновых кислот
Биосинтез пиримидиновых нуклеотидов
Механизм синтеза пиримидиновых нуклеотидов почти полностью расшифрован благодаря исследованиям П. Рейхарда. Показано, что в клетках животных и в микроорганизмах конечными продуктами синтеза также не являются свободные пиримидиновые основания и остаток рибозы присоединяется к уже сформировавшемуся пиримидиновому кольцу. Синтез начинается с элементарных уровней (СО2, NH3, аспартат), и специфическую ключевую роль выполняет оротовая кислота.
Последовательность химических реакций синтеза пиримидиновых нуклеотидов, в частности УМФ, можно представить в следующем виде:
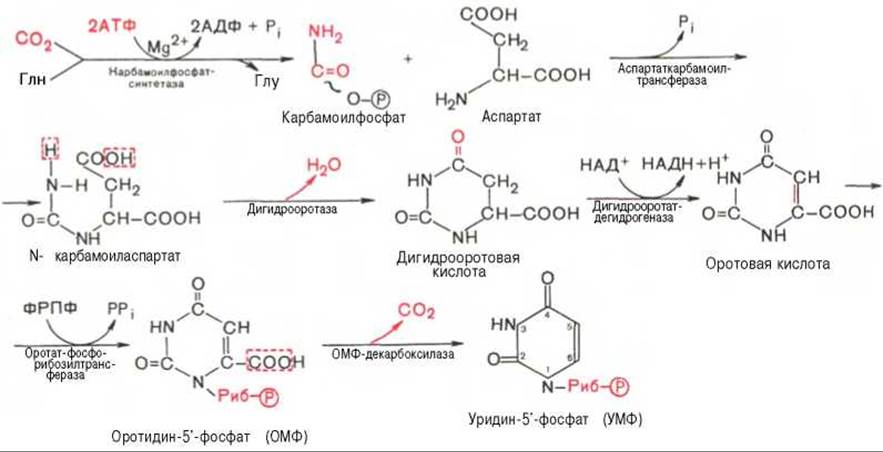
Как видно, I стадия синтеза УМФ включает катализируемое цитоплазматической карбамоилфосфатсинтетазой образование карбамоилфосфата из глутамина (см. главу 12).
На II стадии карбамоилфосфат реагирует с аспартатом, в результате чего образуется N-карбамоиласпарагиновая кислота. Последняя подвергается циклизации (под действием дигидрооротазы) с отщеплением молекулы воды, при этом образуется дигидрооротовая кислота, которая, подвергаясь дегидрированию, превращается в оротовую кислоту. В этой реакции участвует специфический НАД-содержащий фермент дигидрооротатдегидрогеназа. Оротовая кислота обратимо реагирует с ФРПФ, являющимся донатором рибозофосфата, с образованием оротидин-5'-фосфата (ОМФ). Декарбоксилирование последнего приводит к образованию первого пиримидинового нуклеотида — уридин-5-фосфата (УМФ).
Превращение УМФ в УДФ и УТФ осуществляется, как и пуриновых нуклеотидов, путем фосфотрансферазных реакций:
УМФ + АТФ <=> УДФ + АДФ;
УДФ + АТФ <=>УТФ + АДФ.
Биосинтез цитидиловых нуклеотидов. Предшественником цитидиловых нуклеотидов является УТФ, который превращается в ЦТФ:
![]()
У прокариот в этой реакции используется преимущественно свободный аммиак, в то время как в клетках животных ЦТФ-синтетаза катализирует включение амидной группы глутамина в 4-е положение пиримидинового кольца УТФ. Следует отметить, что образующийся ЦТФ служит отрицательным эффектором регуляторного аллостерического фермента аспартаткарбамоилтрансферазы, ингибируя по типу обратной связи начальную стадию биосинтеза пиридиновых нуклеотидов. АТФ предотвращает это ингибирование.
Биосинтез тимидиловых нуклеотидов. Тимидиловые нуклеотиды входят в состав ДНК, содержащей дезоксирибозу. Поэтому сначала рассмотрим механизмы синтеза дезоксирибонуклеотидов. При помощи метода меченых атомов было показано, что этот синтез начинается не со свободной дезоксирибозы, а путем прямого восстановления рибонуклеотидов у 2'-го атома углерода. При инкубации меченых предшественников (рибонуклеотидов) в бесклеточной системе бактерий метку обнаружили в составе дезоксирибонуклеотидов. По данным П. Рейхарда, у Е. coli все 4 рибонуклеозиддифосфата восстанавливаются в соответствующие дезоксианалоги: dАДФ, dГДФ, dЦДФ, dУДФ — при участии сложной ферментной системы, состоящей по меньшей мере из четырех разных ферментов.
Химический смысл превращения рибонуклеотидов в дезоксирибонуклеотиды сводится к элементарному акту —восстановлению рибозы в 2-дезоксирибозу, требующему наличия двух атомов водорода. Непосредственным источником последних оказался восстановленный термостабильный белок тиоредоксин, содержащий две свободные SH-группы на 108 аминокислотных остатков. Тиоредоксин легко окисляется, превращаясь в дисульфидную S-S-форму. Для его восстановления в системе имеется специфический ФАД-содержащий фермент тиоредоксинредуктаза (мол. масса 68000), требующая наличия восстановленного НАДФН. Обозначив условно рибонуклеозиддифосфат РДФ, образование дезоксирибонуклеотидов можно представить следующим образом:

Обе стадии могут быть представлены в виде схемы:

Для синтеза тимидиловых нуклеотидов, помимо дезоксирибозы, требуется также метилированное производное урацила — тимин. Оказалось, что в клетках имеется особый фермент тимидилатсинтаза, катализирующая метилирование не свободного урацила, а dУМФ; реакция протекает по уравнению:
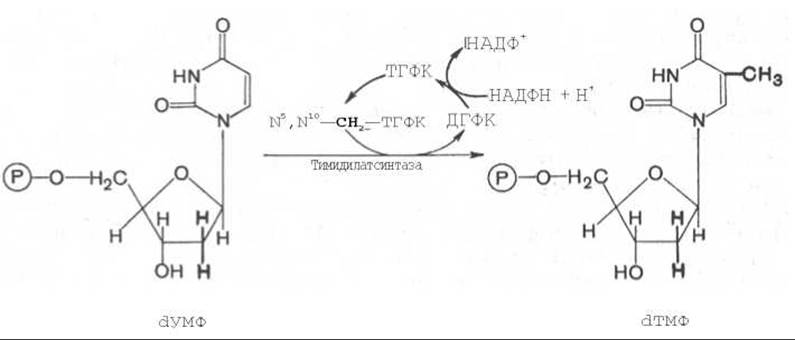
Донором метильной группы в тимидилатсинтазной реакции является N5,N10-метилен-ТГФК, которая одновременно отдает и водородный протон, поэтому одним из конечных продуктов реакции является не тетрагидро-, а дигидрофолиевая кислота (ДГФК). Последняя вновь восстанавливается до ТГФК под действием НАДФН-зависимой дигидрофолатредуктазы. Из образовавшегося ТМФ путем фосфотрансферазных реакций образуются dТДФ и dТТФ.
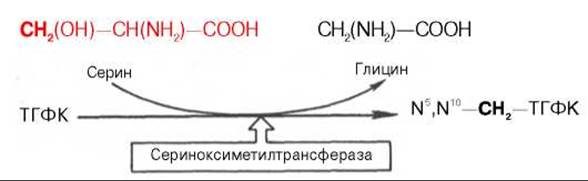
Регенерация N5,N10—СН2—ТГФК, собственно ее биосинтез, представляет определенный интерес. Оказалось, что этот синтез требует участия аминокислоты серина (донатор метильной группы) и пиридоксальфосфатсодержащего фермента сериноксиметилтрансферазы в соответствии с уравнением:
Синтез всех остальных дезоксирибонуклеозид-5'-трифосфатов, непосредственно участвующих в синтезе ДНК, также осуществляется путем фосфорилирования дезоксирибонуклеозид-5'-дифосфатов в присутствии АТФ:

Далее на двух схемах суммированы данные о взаимопревращениях пуриновых и пиримидиновых нуклеотидов, а также о связи их с синтезом нуклеиновых кислот. Как видно из схем, в образовании пуриновых и пиримидиновых нуклеотидов специфическое участие принимает ФРПФ, являющийся донором фосфорибозильного остатка в биосинтезе как оротидин-5'-фосфата, так и ИМФ; последние считаются ключевыми субстратами в синтезе нуклеиновых кислот в клетках.

