Биологическая химия - Березов Т. Т., Коровкин Б. Ф. 1998
Химия нуклеиновых кислот
Структура нуклеиновых кислот
Для понимания ряда особенностей структуры ДНК важное значение имели закономерности состава и количественного содержания азотистых оснований, установленные впервые Э. Чаргаффом. Оказалось, что азотистые основания ДНК обычно варьируют у разных видов организмов, однако почти не претерпевают изменений у одного и того же вида в процессе развития или в зависимости от изменений окружающей среды либо характера питания. Показано также, что ДНК, выделенная из разных тканей одного и того же вида, имеет одинаковый состав азотистых оснований. Полученные количественные соотношения были названы правилами Чаргаффа. При анализе состава очищенной ДНК, выделенной из разных источников, были сделаны следующие выводы:
1) молярная доля пуринов* равна молярной доле пиримидинов:
![]()
2) количество аденина и цитозина равно количеству гуанина и тимина:
![]()
3) количество аденина равно количеству тимина, а количество гуанина равно количеству цитозина: А = Т и Г = Ц; соответственно
![]()
4) существенным для характеристики вида (таксономическое значение) оказался так называемый коэффициент специфичности, отражающий отношение
![]()
* Здесь и далее пуриновые и пиримидиновые основания обозначены начальными буквами соответствующего названия.
Это отношение часто выражают в молярных процентах (Г + Ц), или процентах ГЦ-пар. Для животных и большинства растений этот коэффициент ниже 1 (от 0,54 до 0,94), у микроорганизмов он колеблется в значительных пределах (от 0,45 до 2,57).
Данные, полученные А. Н. Белозерским и его учениками, свидетельствуют о существовании в природе АТ-типа ДНК (у хордовых и беспозвоночных животных, высших растений, ряда бактерий, дрожжевидных организмов) и ГЦ-типа ДНК (у недрожжевидных грибов, актиномицетов, ряда бактерий и вирусов).
Известно, что структурными единицами нуклеиновых кислот являются мономерные молекулы — мононуклеотиды. Следовательно, нуклеиновые кислоты представляют собой полинуклеотиды. Это продукты полимеризации мононуклеотидов, число и последовательность расположения которых в цепях дНк и РНК определяются в строгом соответствии с программой, заложенной в молекуле матрицы (см. главу 14). Мононуклеотиды легко образуются при гидролизе ДНК и РНК в присутствии нуклеаз, состоят из трех специфических компонентов: азотистого основания, углевода и фосфорной кислоты. В этой «триаде» мононуклеотида углевод занимает среднее положение. Соединения азотистого (любого) основания и углевода (рибозы или дезоксирибозы), получившие название нуклеозидов, легко образуются из мононуклеотида при гидролитическом отщеплении фосфорной кислоты в присутствии щелочи или при участии специфических ферментов — нуклеотидаз.
Нуклеозиды содержат пуриновое или пиримидиновое основание, соединенное с углеводом N-гликозидной связью. В составе нуклеиновых кислот обнаруживаются только ß-нуклеозиды. Примером могут служить два мононуклеотида: аденозин-5'-монофосфорная кислота (АМФ) и цитидин-5'-монофосфорная кислота (ЦМФ):
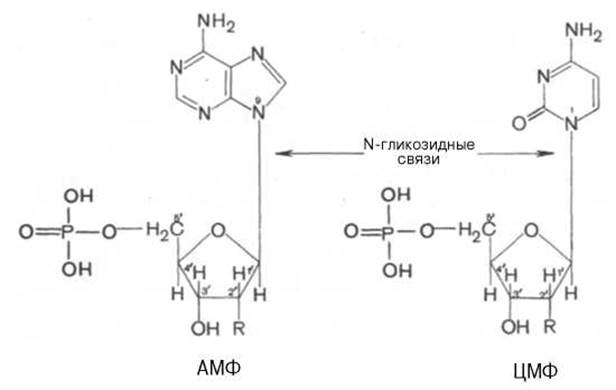
R у 2' углерода представлен Н- или ОН-группой в зависимости от типа нуклеиновой кислоты — ДНК или РНК. В образовании N-гликозидной связи в пуриновых нуклеотидах принимают участие N-9 пурина и С-1' пентозы, а в пиримидиновых нуклеотидах — N-1 пиримидина и С-1' пентозы. Чтобы отличить углеродные атомы рибозы или дезоксирибозы от углеродных атомов, входящих в состав пуриновых и пиримидиновых оснований, первые принято обозначать символом «штрих»: например, атомы у 3-го и 5-го углерода обозначают С-3' и С-5' или, чаще, 3' и 5'.
Следует отметить, что среди продуктов ферментативного гидролиза ДНК и РНК обнаруживаются, помимо нуклеозид-5'-монофосфатов, также нуклеозид-3'-монофосфаты. Положение фосфата определяется местом разрыва фосфодиэфирной связи между соседними нуклеотидами, что указывает на характер связи нуклеотидов через остаток фосфорной кислоты, соединяющий 3' и 5' углеродные атомы пентозы.
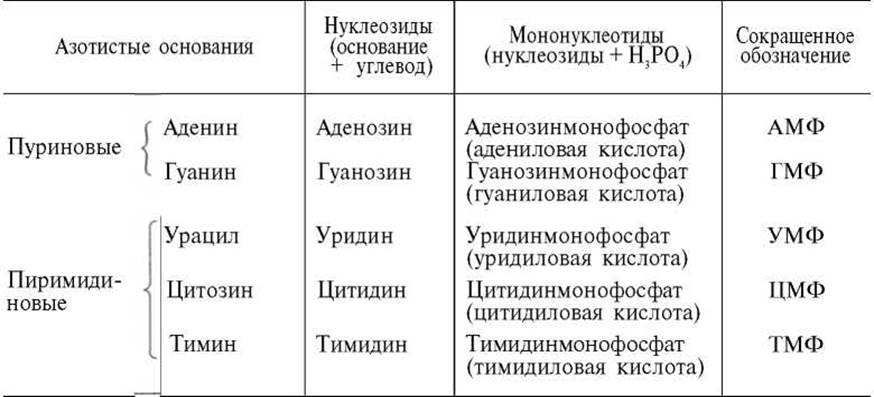
В табл. 3.3 приведены состав и названия (включая тривиальные), а также сокращенные обозначения нуклеозидов и нуклеотидов (для РНК они называются рибонуклеотидами, а для ДНК — дезоксирибонуклеотидами).
Таблица 3.3. Состав нуклеозидов и мононуклеотидов
Мононуклеотиды и их производные, а также динуклеотиды присутствуют в клетках в свободном виде и играют важную роль в обмене веществ. В частности, нуклеотидную структуру имеют многие коферменты, включая коферменты оксидоредуктаз. Мононуклеотиды, присоединяя еще один остаток фосфата, образуют фосфоангидридную связь (наподобие связи, имеющейся в пирофосфате) и превращаются в нуклеозиддифосфаты (соответственно они обозначаются сокращенно АДФ, ГДФ, УДФ, ЦДФ и ТДФ). Последние, присоединяя еще один остаток фосфата, образуют нуклеозидтрифосфаты (соответственно обозначаются АТФ, ГТФ, УТФ, ЦТФ и ТТФ).
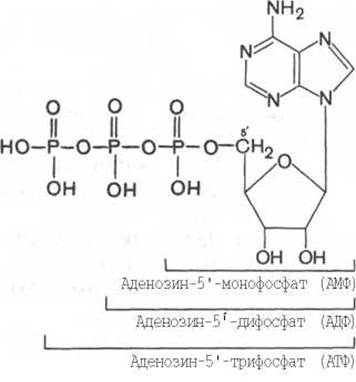
Следует особо указать, что только свободные нуклеозидтрифосфаты в клетках являются предшественниками ферментативного синтеза ДНК и РНК (см. главу 13). Однако в клетках имеются свободные, также природные нуклеозидтрифосфаты, не принимающие участия в синтезе белка, но выполняющие жизненно важные функции. В частности, одной из важнейших функций нуклеозидтрифосфатов и особенно АТФ является их участие в биоэнергетике всех живых организмов (см. главу 13). Приводим схему образования молекул аденозинди- и аденозинтрифосфатов (некоторые атомы водорода, как и углерода, в пуриновом ядре и в кольце рибозы опущены):
Необходимо указать на существование в организме еще двух типов фосфорных эфиров нуклеотидов, когда фосфат связывает 2 атома кислорода пентозного остатка в одном и том же нуклеотиде и когда фосфатный мостик объединяет два разных мононуклеотида. Примером первого типа являются циклические нуклеотиды 2',3'- и 3',5'-, т.е. два возможных класса соединений, в которых кислородные атомы у С-2' и С-3' или у С-3' и С-5' участвуют в образовании циклической структуры:
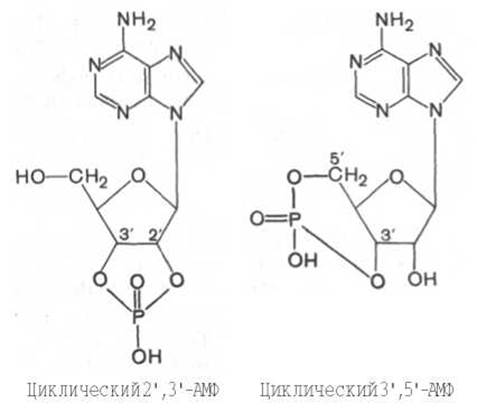
Первое из этих соединений, 2',3'-АМФ, образуется в качестве промежуточного продукта распада рибонуклеиновых кислот, в то время как циклический 3',5'-АМФ (цАМФ) является естественно встречающимся рибонуклеотидом (он образуется из АТФ в процессе реакции, катализируемой ферментом аденилатциклазой). цАМФ наделен рядом уникальных функций и высокой биологической активностью в регуляции процессов обмена, выполняя роль медиатора внеклеточных сигналов в клетках животных. Аналогичной функцией наделены цГМФ, производные УДФ, ЦТФ и нуклеотиды в составе кофакторов и коферментов (см. главу 4).
Примером, когда одна фосфатная группа связывает два разных рибонуклеотида, является структура гуанозилтимидинфосфата:
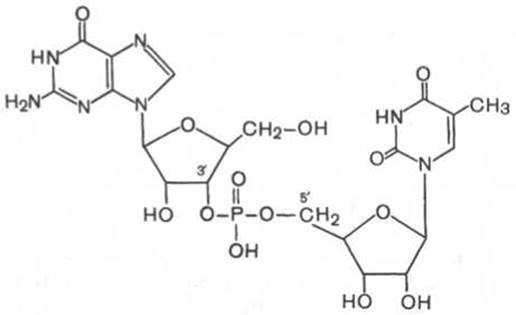
Это соединение хотя и не обнаружено в природе, но содержит тип связи, характерный для природных нуклеиновых кислот. Видно, что С-3' рибозы одного нуклеотида соединяется посредством фосфодиэфирной связи с С-5' рибозы второго нуклеотида.
В медицинской практике, в частности в онкологии, нашли широкое применение синтетические аналоги как азотистых оснований, так нуклеозидов и нуклеотидов. Эти аналоги, имеющие небольшие модификации в структуре основания или углевода, встраиваясь в соответствующие клеточные компоненты, оказывают заметный цитотоксический эффект. К наиболее распространенным лекарственным препаратам — аналогам пуриновых и пиримидиновых оснований (и соответствующим нуклеотидам) относятся 5-фторурацил, 6-тио- и 6-меркаптопурин, 8-азагуанин, 6-азауридин и 6-азацитидин, а также 5-йодпроизводное дезоксиуридина.
Помимо сокращенных названий и обозначений нуклеозидов и нуклеотидов (см. табл. 3.3), приняты буквенные обозначения нуклеозидов (и нуклеотидов): в частности, для аденозина (и АМФ) это А, для гуанозина (и ГМФ) — Г, для цитидина (и ЦМФ) — Ц, для уридина (и УМФ)—У, для тимидина (и ТМФ) — Т. Пользуясь этими символами, приведенный выше дирибонуклеозидмонофосфат можно обозначить как Г—Т. Заметим, что как по структуре, так и по свойствам Г—Т и Т—Г будут сильно отличаться друг от друга (как и в случае дипептидов).