Биологическая химия - Березов Т. Т., Коровкин Б. Ф. 1998
Химия нуклеиновых кислот
Структура нуклеиновых кислот
Вторичная структура нуклеиновых кислот
В соответствии с моделью Дж. Уотсона и Ф. Крика, предложенной в 1953 г. на основании ряда аналитических данных, а также рентгеноструктурного анализа молекула ДНК состоит из двух цепей, образуя правовращающую спираль, в которую обе полинуклеотидные цепи закручены вокруг одной и той же оси. Удерживаются цепи благодаря водородным связям, образующимся между их азотистыми основаниями (рис. 3.1). Обе цепи полинуклеотидов в биспиральной молекуле ДНК имеют строго определенное пространственное расположение, при котором азотистые основания находятся внутри, а фосфорильные и углеводные компоненты — снаружи*.
Детальный анализ всевозможных вариантов образования водородных связей между основаниями показал, что в биспиральной молекуле ДНК основания уложены парами: пурин из одной цепи и пиримидин из другой в соответствии с правилами Чаргаффа. Поскольку ориентация оснований на плоскости не является, очевидно, произвольной, и основания в полинуклеотидах представлены в лактамной форме, наиболее вероятными были признаны пары аденин — тимин и гуанин-цитозин. Этот способ спаривания получил в дальнейшем экспериментальное подтверждение. Избирательность взаимодействия пар А—Т и Г—Ц принято выражать термином «комплементарность», а соответствующие азотистые основания называют комплементарными. Стабильность А—Т оснований обеспечивается двумя водородными связями, а пар Г—Ц — тремя, что в свою очередь определяется особенностями расположения функциональных групп азотистых оснований. Длина водородных связей между основаниями составляет около 0,3 нм. Таким образом, комплементарными оказываются не только отдельные основания, но и дезоксирибонуклеотидные цепи ДНК в целом, способствующие образованию весьма компактной структуры и стабилизации всей молекулы*.
* За это открытие Дж. Уотсон и Ф. Крик вместе с М. Уилкинсом получили Нобелевскую премию в 1962 г.
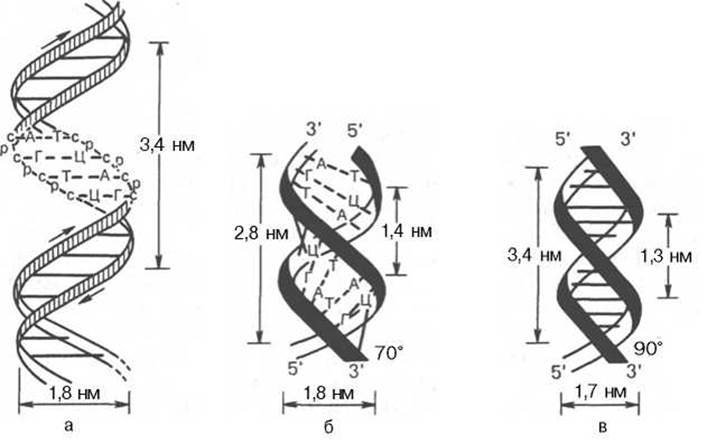
Рис. 3.1. Схематическое изображение двойной спирали ДНК.
а - по Уотсону и Крику: с - остаток дезоксирибозы, р - остаток фосфорной кислоты; б - А-форма ДНК; в - В-форма ДНК.
Обе цепи в молекуле ДНК имеют противоположную полярность. Это означает, что межнуклеотидная связь в одной цепи имеет направление 5’→3’, а в другой — 3'→5'. Подобная направленность цепей имеет важное биологическое значение при репликации и транскрипции молекулы ДНК.
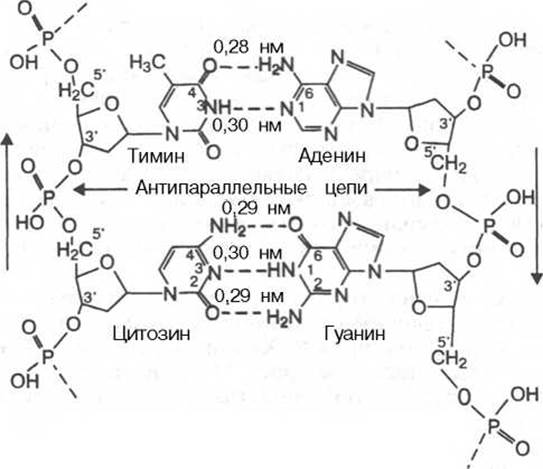
* Данные, полученные в последние годы, свидетельствуют, что в стабилизации биспиральной структуры основную роль играют гидрофобные взаимодействия между комплементарными основаниями, стыкующимися в центре двойной спирали. Водородные связи, вероятнее всего, обеспечивают специфичность спаривания оснований.
На модели ДНК (см. рис. 3.1) видно, что расстояние между витками (шаг спирали) равно 3,4 нм. На этом участке укладываются 10 нуклеотидных остатков, размер одного нуклеотида составляет 0,34 нм; диаметр биспиральной молекулы равен 1,8 нм.
Необходимо указать, что конфигурация двойной спирали ДНК сильно меняется в зависимости от количественного содержания воды и ионной силы окружающей среды. Методами рентгеноструктурного анализа доказано существование по крайней мере 6 форм ДНК, названных А-, В-, С-, D-, Е- и Z-формами. Конфигурация двух из них в простейшей форме представлена на рис. 3.1, б и в. Можно увидеть, что у А-формы наблюдается некоторое смещение пар оснований от оси молекулы к периферии, что отражается на размерах (2,8 нм — длина одного витка, в котором вместо 10 содержится 11 мононуклеотидов; меняется расстояние между нуклеотидами и др.). Если А- и В-формы представляют собой правозакрученную двойную спираль, то Z-форма (зигзагообразная) ДНК имеет левозакрученную конфигурацию, в которой фосфодиэфирный остов располагается зигзагообразно вдоль оси молекулы. Параллельно фосфодиэфирному остову в структуре А- и В-форм ДНК имеются большая и малая бороздки (желобки) — сайты, где присоединяются белки, выполняющие, очевидно, регуляторные функции при экспрессии генов. В настоящее время есть основание считать, что между А- и В-формами ДНК осуществляются взаимные переходы при изменении концентрации соли и степени гидратации. В-форма ДНК больше всего подходит к модели Уотсона и Крика. В этих переходах, которые могут быть вызваны растворителями или белками, очевидно, заключен определенный биологический смысл. Предполагают, что в А-форме ДНК выполняет роль матрицы в процессе транскрипции (синтез РНК на молекуле ДНК), а в В-форме — роль матрицы в процессе репликации (синтез ДНК на молекуле ДНК).
В структуре ДНК, как и в структуре РНК, открыты нуклеотидные последовательности, получившие название «палиндромы», или перевернутые повторы. Они встречаются как внутри одной цепи, так и в двойной спирали. Например, как слово ротатор, которое одинаково читается как справа налево, так и обратно. Подобные обратные повторы могут служить основой для образования структуры шпилек или других вариаций с измененным внутрицепочечным и межцепочечным спариванием и формированием на отдельных участках тройной спирали. Возможно, эти палиндромные структуры имеют определенный биологический смысл в регуляции экспрессии отдельных генов, выполняя роль сайтов для ДНК-связывающих белков. Предстоит, однако, приложить немало усилий для установления как точной структуры этих вариаций, так и для определения их функциональной роли.
Менее охарактеризована вторичная структура матричных и рибосомных РНК. Относительно вторичной структуры тРНК наиболее вероятной представляется модель, предложенная Р. Холли, плоское изображение которой напоминает клеверный лист (см. рис. 14.3). В настоящее время, когда известна первичная структура большинства тРНК, последовательность всех или почти всех природных тРНК как будто бы укладывается в эту схему «клеверного листа» (см. главу 14). При сравнении этих структур выявляется ряд закономерностей, несомненно, имеющих определенный биологический смысл. Во всех тРНК есть участки, взаимодействующие с рибосомами, места для связывания с аминокислотами и ферментами, а также специфическая последовательность трех нуклеотидов (триплет), называемая антикодоном, которая оказывается комплементарной тринуклеотидной последовательности мРНК (кодону), кодирующей включение в белковую молекулу определенной аминокислоты.
Независимо от типа РНК синтезированный в клетке продукт транскрипции (см. главу 13) всегда представлен единственной цепью, упакованной во вторичную структуру не случайно, а в соответствии с программой ДНК. Поскольку в составе РНК имеются свободные 2'-оксигруппы рибозы, не связанные со стандартным крик-уотсоновским спариванием азотистых оснований, появляются дополнительные возможности образования вторичной и третичной структур, содержащих выпуклости, шпильки, или крестообразные структуры. Особенности структуры тРНК имеют прямое отношение к процессу трансляции, поэтому более подробно они рассмотрены в разделе биосинтеза белка (глава 14).