Биологическая химия - Березов Т. Т., Коровкин Б. Ф. 1998
Ферменты
Основные свойства ферментов
К ферментам применимы три основных критерия, характерных и для неорганических катализаторов. В частности, они остаются неизмененными после реакции*, т.е. освобождаясь, могут вновь реагировать с новыми молекулами субстрата (хотя нельзя исключить побочных влияний условий среды на активность фермента). Ферменты способны оказывать действие в ничтожно малых концентрациях (например, одна молекула фермента реннина, содержащегося в слизистой оболочке желудка теленка, створаживает около 106 молекул казеиногена молока за 10 мин при температуре 37°С). Наличие либо отсутствие фермента или любого другого катализатора не оказывает влияния на величину константы равновесия и свободной энергии (∆G). Катализаторы лишь повышают скорость, с которой система приближается к термодинамическому равновесию, не сдвигая точки равновесия. Химические реакции с высокой константой равновесия и отрицательной величиной AG принято называть экзергоническими. Реакции с низкой константой равновесия и соответственно положительной величиной ∆G (они обычно не протекают спонтанно) называются эндергоническими. Для начала и завершения этих реакций необходим приток энергии извне. В живых системах экзергонические процессы обычно сопряжены с эндергоническими реакциями, обеспечивая последние необходимым количеством энергии.
* Доказано, что некоторые ферменты в процессе химической реакции подвергаются модификации и даже распаду под действием конечных продуктов реакции, а не освобождаются в неизмененном виде, как утверждал Л. Михаэлис; типичный пример — цитохромы класса Р450.
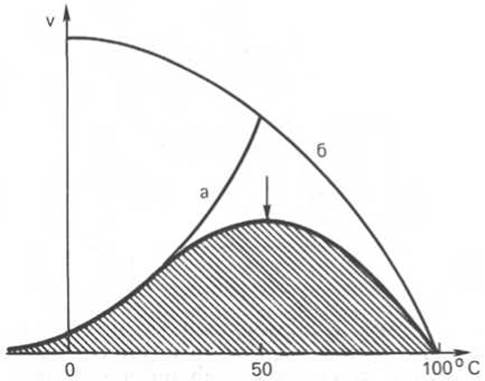
Рис. 4.16. Зависимость скорости катализируемой ферментом реакции от температуры.
а - повышение скорости реакции как функция температуры; б - снижение скорости реакции как функция денатурации белка-фермента; стрелка указывает оптимум температуры.
Ферменты, являясь белками, обладают рядом характерных для этого класса органических соединений свойств, отличающихся от свойств неорганических катализаторов.
Термолабильность ферментов. Скорость химических реакций зависит от температуры, поэтому катализируемые ферментами реакции также чувствительны к изменениям температуры. Установлено, что скорость большинства биохимических реакций повышается в 2 раза при повышении температуры на 10°С и, наоборот, снижается в 2 раза при понижении температуры на 10°С. Этот показатель получил название температурного коэффициента. Однако вследствие белковой природы фермента тепловая денатурация при повышении температуры будет снижать эффективную концентрацию фермента с соответствующим снижением скорости реакции. Так, при температуре, не превышающей 45—50°С, скорость реакции увеличивается согласно теории химической кинетики. При температуре выше 50° С на скорость реакции большое влияние начинает оказывать тепловая денатурация белка-фермента, приводящая к полному прекращению ферментативного процесса (рис. 4.16).
Таким образом, термолабильность, или чувствительность к повышению температуры, является одним из характерных свойств ферментов, резко отличающих их от неорганических катализаторов. В присутствии последних скорость реакции возрастает экспоненциально при повышении температуры (см. кривую «а» на рис. 4.16). При температуре 100°С почти все ферменты утрачивают свою активность (исключение составляет, очевидно, только один фермент мышечной ткани — миокиназа, которая выдерживает нагревание до 100°С). Оптимальной для действия большинства ферментов теплокровных животных является температура 40°С; в этих условиях скорость реакции оказывается максимальной вследствие увеличения кинетической энергии реагирующих молекул. При низких температурах (0°С и ниже) ферменты, как правило, не разрушаются, хотя активность их падает почти до нуля. Во всех случаях имеет значение время воздействия соответствующей температуры. В настоящее время для пепсина, трипсина и ряда других ферментов доказано существование прямой зависимости между скоростью инактивации фермента и степенью денатурации белка. Следует отметить, что на термолабильность ферментов определенное влияние оказывает концентрация субстрата, рН среды и другие факторы.
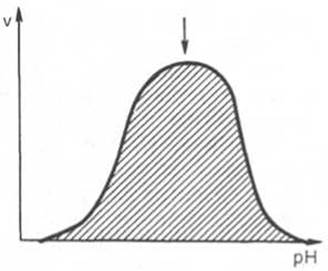
Рис. 4.17. Зависимость скорости катализируемой ферментом реакции от рН (стрелка указывает оптимум рН).
Зависимость активности ферментов от рН среды. Ферменты обычно наиболее активны в пределах узкой зоны концентрации водородных ионов, соответствующей для животных тканей в основном выработанным в процессе эволюции физиологическим значениям рН среды 6,0—8,0. При графическом изображении на кривой колоколообразной формы имеется определенная точка, в которой фермент проявляет максимальную активность; эту точку называют оптимумом рН среды для действия данного фермента (рис. 4.17). При определении зависимости активности фермента от концентрации водородных ионов реакцию проводят при разных значениях рН среды, обычно при оптимальной температуре и наличии достаточно высоких (насыщающих) концентраций субстрата. В табл. 4.3 приводятся оптимальные значения рН среды для ряда ферментов.
Таблица 4.3. Оптимальные значения рН для некоторых ферментов
Фермент |
рН |
Фермент |
рН |
Пепсин |
1,5—2,5 |
Каталаза |
6,8—7,0 |
Катепсин В |
4,5—5,0 |
Уреаза |
7,0—7,2 |
Амилаза из солода |
4,9—5,2 |
Липаза |
7,0—8,5 |
панкреатическая |
|||
Сахараза кишечная |
5,8—6,2 |
Трипсин |
7,5—8,5 |
Амилаза слюны |
6,8—7,0 |
Аргиназа |
9,5-10,0 |
Из данных табл. 4.3 видно, что рН-оптимум действия ферментов лежит в пределах физиологических значений. Исключение составляют пепсин, рН-оптимум которого 2,0 (при рН 6,0 он не активен и не стабилен). Объясняется это, во-первых, структурной организацией молекулы фермента и, во-вторых, тем, что пепсин является компонентом желудочного сока, содержащего свободную соляную кислоту, которая создает оптимальную кислую среду для действия этого фермента. С другой стороны, рН-оптимум аргиназы лежит в сильнощелочной зоне (около 10,0); такой среды нет в клетках печени, следовательно, in vivo аргиназа функционирует, по-видимому, не в своей оптимальной зоне рН среды.
Согласно современным представлениям, влияние изменений рН среды на молекулу фермента заключается в воздействии на состояние и степень ионизации кислотных и основных групп (в частности, СООН-группы дикарбоновых аминокислот, SH-группы цистеина, имидазольного азота гистидина, NH2-группы лизина и др.). При резких сдвигах от оптимума рН среды ферменты могут подвергаться конформационным изменениям, приводящим к потере активности вследствие денатурации или изменения заряда молекулы фермента. При разных значениях рН среды активный центр может находиться в частично ионизированной или неионизированной форме, что сказывается на третичной структуре белка и соответственно на формировании активного фермент-субстратного комплекса. Имеет значение, кроме того, состояние ионизации субстратов и кофакторов.
Специфичность ферментов. Ферменты обладают высокой специфичностью действия. Это свойство часто существенно отличает их от неорганических катализаторов. Так, мелкоизмельченные платина и палладий могут катализировать восстановление (с участием молекулярного водорода) десятков тысяч химических соединений различной структуры. Высокая специфичность ферментов обусловлена, как было отмечено, конформационной и электростатической комплементарностью между молекулами субстрата и фермента и уникальной структурной организацией активного центра, обеспечивающими «узнавание», высокое сродство и избирательность протекания одной какой-либо реакции из тысячи других химических реакций, осуществляющихся одновременно в живых клетках.
В зависимости от механизма действия различают ферменты с относительной (или групповой) и абсолютной специфичностью. Так, для действия некоторых гидролитических ферментов наибольшее значение имеет тип химической связи в молекуле субстрата. Например, пепсин в одинаковой степени расщепляет белки животного и растительного происхождения, несмотря на то что эти белки существенно отличаются друг от друга как по химическому строению и аминокислотному составу, так и по физико-химическим свойствам. Однако пепсин не расщепляет ни углеводы, ни жиры. Объясняется это тем, что точкой приложения, местом действия пепсина является пептидная —СО—NH-связь. Для действия липазы, катализирующей гидролиз жиров на глицерин и жирные кислоты, подобным местом является сложноэфирная связь. Аналогичной групповой специфичностью обладают трипсин, химотрипсин, пептидазы, ферменты, гидролизующие а-гликозидные связи (но не ß-гликозидные связи, имеющиеся в целлюлозе) в полисахаридах, и др. Обычно эти ферменты участвуют в процессе пищеварения, и их групповая специфичность, вероятнее всего, имеет определенный биологический смысл. Относительной специфичностью наделены также некоторые внутриклеточные ферменты, например гексокиназа, катализирующая в присутствии АТФ фосфорилирование почти всех гексоз, хотя одновременно в клетках имеются и специфические для каждой гексозы ферменты, выполняющие такое же фосфорилирование (см. главу 10).
Абсолютной специфичностью действия называют способность фермента катализировать превращение только единственного субстрата. Любые изменения (модификации) в структуре субстрата делают его недоступным для действия фермента. Примерами таких ферментов могут служить аргиназа, расщепляющая в естественных условиях (в организме) аргинин, уреаза, катализирующая распад мочевины, и др.
Имеются экспериментальные доказательства существования так называемой стереохимической специфичности, обусловленной существованием оптически изомерных L- и D-форм или геометрических (цис- и транс-) изомеров химических веществ. Так, известны оксидазы L-и D -аминокислот, хотя в природных белках обнаружены только L-аминокислоты. Каждый из видов оксидаз действует только на свой специфический стереоизомер*.

Наглядным примером стереохимической специфичности является бактериальная аспартатдекарбоксилаза, катализирующая отщепление СО2 только от L-аспарагиновой кислоты с превращением ее в L-аланин. Стереоспецифичность проявляют ферменты, катализирующие и синтетические реакции. Так, из аммиака и а-кетоглутарата во всех живых организмах синтезируется L-изомер глутаминовой кислоты, входящей в состав природных белков. Если какое-либо соединение существует в форме цис- и транс-изомеров с различным расположением групп атомов вокруг двойной связи, то, как правило, только один из этих геометрических изомеров может служить в качестве субстрата для действия фермента. Например, фумараза катализирует превращение только фумаровой кислоты (трансизомер), но не действует на малеиновую кислоту (цис-изомер):

Таким образом, благодаря высокой специфичности действия ферменты обеспечивают протекание с большой скоростью лишь определенных химических реакций из огромного разнообразия возможных превращений в микропространстве клеток и целостном организме, регулируя тем самым интенсивность обмена веществ.