БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ - Е. С. Северина - 2004
РАЗДЕЛ 16. ОНКОГЕНЕЗ
VI. Инвазия и метастазирование
Доброкачественные опухоли иногда могут расти быстро и достигать больших размеров, но они не метастазируют. Только для злокачественных опухолей обнаруживается способность прорастать в другие ткани, как соседние, так и отдалённые, где они образуют вторичные опухоли.
Первоначально опухолевые клетки образуют клон генетически идентичных (моноклональных) клеток, которые делятся чаще, чем соседние, нормальные клетки. Они не запрограммированы к движению. Однако состав и поведение таких клеток не статичны, и потомки одной клетки начинают «расходиться» как генетически, так и фенотипически. Каждое последующее поколение клеток обнаруживает увеличение отклонений от нормы.
Когда клеточная масса опухоли достигает диаметра около 2 мм, клетки секретируют белковые факторы, стимулирующие рост соединительной ткани, которая бы окружила опухоль, и васкулярные клетки, индуцирующие рост кровеносных сосудов, или ангиогенез. Установлено, что рост сосудов является ключевым звеном в прогрессии опухоли. Щелочной и кислый факторы роста фибробластов, секретируемые опухолевыми клетками, стимулируют пролиферацию эндотелиальных клеток и образование новых капилляров. Ангиогенез создаёт опухолевым клеткам дополнительные преимущества для роста и инвазии.
А. Изменение состава мембранных белков
В метастазирующих клетках происходит существенное изменение состава мембранных белков. В эпителиальных клетках основным белком, ответственным за адгезивные свойства, служит Е-кадгерин (см. раздел 5), внеклеточный домен которого ответственен за образование межклеточных связей с молекулами Е-кадгерина соседних клеток. Другой класс белков — катенины — отвечает за связывание Е-кадгерина с цитоскелетом.
В опухолевых клетках содержание Е-кадгерина снижено, а молекулы катенинов либо функционально неактивны, либо отсутствуют. Мутации в гене Е-кадгерина редки, но если они возникают, то у таких пациентов наблюдают низкодифференцированные формы рака. Контактное торможение и связь клеток друг с другом нарушаются, внутриклеточная архитектоника мало напоминает структуру нормальных клеток.
Было доказано, что (3-катенин участвует также во внутриклеточной системе передачи сигналов и стимулирует пролиферацию клеток. Однако, будучи в комплексе с АРС-белком (от англ. adenomatous poliposis coli), он ингибирует деление и участвует в уничтожении дефектных клеток через апоптоз. Если в результате мутаций один из этих белков будет изменён, то комплекс не образуется, и β-катенин, не сдерживаемый АРС-белком, взаимодействует с ДНК и стимулирует вступление клеток в клеточный цикл. Появление мутаций в АРС коррелирует с ростом полипов (небольших доброкачественных опухолей) в прямой кишке, а у людей с наследственной формой мутантного АРС повышается риск перерождения полипов в опухоль.
В связывании клеток с коллагеном участвуют белки-интегрины, а с другими компонентами межклеточного матрикса и базальными мембранами — фибронектин и ламинины (см. раздел 15). При трансформации клеток количественное и качественное содержание этих белков меняется. В большинстве опухолей снижено количество фибронектина и синтезируются модифицированные интегрины, которые помогают инвазивным клеткам мигрировать через соединительную ткань и стенку капилляров.
Б. Ферменты, приспосабливающие опухолевые клетки к движению
Инвазия — активный процесс, включающий стадии, в которых опухолевая клетка:
✵ проходит через межклеточный матрикс, достигая кровеносного или лимфатического сосуда;
✵ преодолевает стенку сосуда и поступает в кровеносное русло или лимфу;
✵ циркулирует с током крови в виде надмолекулярных комплексов с белками и клетками крови;
✵ прикрепляется к стенке сосуда и повторяет процесс в обратном направлении, продвигаясь на 2 — 3 клеточных диаметра в инвазируемую ткань;
✵ закрепляется и начинает формировать новую опухоль.
Выполнение этих функций требует синтеза специфических ферментов, рецепторов и энергии. Метастазирующие клетки и окружающие опухолевую ткань фибробласты секретируют целый набор ферментов, обеспечивающих разрушение межклеточного матрикса и базальных мембран: коллагеназы, расщепляющие коллаген межклеточного матрикса; гепаразу, катализирующую гидролиз гепаран- сульфата — преобладающего протеогликана базальной мембраны; катепсин В — мощную протеазу, которая в нормальных клетках локализована в лизосомах, а у метастазирующих клеток встроена в плазматическую мембрану и помогает им покинуть родительскую ткань. Этот фермент активирует проколлагеназу, которая специфически расщепляет коллаген IV типа (см. раздел 15); плазмин, который расщепляет некоторые белки межклеточного матрикса неколлагенового происхождения; семейство металлопротеаз, участвующее в разрушении различных компонентов межклеточного матрикса. Они секретируются в виде проферментов и активируются либо катепсином В, либо урокиназой типа активатора плазминогена.
В. Циркуляция метастазирующих клеток
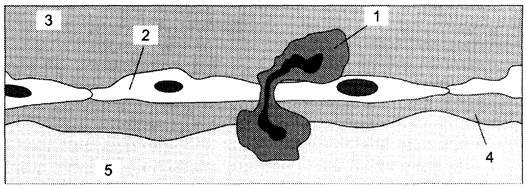
После успешного прохождения через соединительную ткань органа опухолевые клетки продвигаются к ближайшему кровеносному сосуду, проталкиваются между эндотелиальными клетками, выстилающими сосудистую стенку, и выходят в кровоток (рис. 16-15).
Рис. 16-15. Миграция опухолевых клеток в кровеносный или лимфатический сосуд. 1 — делящаяся опухолевая клетка; 2 — неделящаяся опухолевая клетка; 3 — эндотелиальная клетка; 4 — базальная мембрана; 5 — строма; 6 — просвет кровеносного сосуда.

Кровеносные сосуды являются каналами, по которым опухолевые клетки доставляются к местам новой локализации. Они транспортируются по крови в виде комплексов с тромбоцитами, миграционными факторами и фрагментами межклеточного матрикса, которые маскируют их от иммунологического надзора и обеспечивают прикрепление к базальной мембране в органах-мишенях.
Г. Формирование вторичных опухолей
Углеводы, выступающие на поверхность опухолевых клеток, связываются с селектином — углеводным компонентом рецепторов эндотелиальных клеток. Каждый тип опухолевых клеток имеет на плазматической мембране характерную для них углеводную часть, которая может взаимодействовать лишь с определёнными олиго- и полисахаридами клеток эндотелия. Однако эти взаимодействия между углеводами слабы, и клетка окончательно прикрепляется к стенке сосуда с помощью интегринов (рис. 16-16).
Рис. 16-16. Начало формирования вторичной опухоли. 1 — опухолевая клетка; 2 — эндотелиальная клетка; 3 — просвет кровеносного сосуда; 4 — базальная мембрана; 5 — строма.
Эти особенности закрепления опухолевой клетки в другом органе лежат в основе тропности процесса: метастазы появляются не в любых, а в «излюбленных» данной формой опухоли местах. Так, рак простаты, как правило, даёт метастазы в кости, рак молочной железы и лёгкого — в мозг, а рак прямой кишки — в печень.