БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ - Е. С. Северина - 2004
РАЗДЕЛ 4. БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ И БЕЛКОВ (МАТРИЧНЫЕ БИОСИНТЕЗЫ). ОСНОВЫ МОЛЕКУЛЯРНОЙ ГЕНЕТИКИ
Нуклеиновые кислоты — высокомолекулярные соединения со строго определённой линейной последовательностью мономеров. Структура ДНК и РНК — способ «записи информации», обеспечивающий формирование в организме двух информационных потоков. Один из потоков осуществляет воспроизведение информации, заключённой в молекулах ДНК. Удвоение молекул ДНК называют «репликация». В результате этого процесса и последующего деления дочерние клетки наследуют геном родительской клетки, в котором содержится полный набор генов, или «инструкций» о строении РНК и всех белков организма.
Второй поток информации реализуется в процессе жизнедеятельности клетки. В этом случае происходит «считывание», или транскрипция, генов в форме полинуклеотидных последовательностей мРНК и использование их в качестве матриц для синтеза соответствующих белков. В последнем случае осуществляется «перевод» (трансляция)информации, заключённой в мРНК, на «язык» аминокислот. Этот поток информации от ДНК через РНК на белок получил название «центральная догма биологии». Он характерен для всех живых организмов, за исключением некоторых РНК-содержащих вирусов.
Матричная природа синтеза нуклеиновых кислот и белков обеспечивает высокую точность воспроизведения информации. Так, в ходе репликации дочерние молекулы ДНК синтезируются на нитях материнской ДНК. При образовании всех видов РНК, необходимых для синтеза белков, информация об их структуре «считывается» с определённых генов в молекулах ДНК. В синтезе новых молекул белков матрицей, содержащей информацию об их строении, являются мРНК.
Исправление ошибок, возникающих в структуре ДНК под воздействием факторов внешней и внутренней среды, осуществляет ещё один матричный синтез — репарация. Он является вариантом ограниченной репликации и восстанавливает первоначальную структуру ДНК, используя в качестве матрицы участок неповреждённой нити ДНК. При размножении РНК-содержащих вирусов в клетках эукариотических организмов новые молекулы ДНК могут синтезироваться с помощью процесса, в ходе которого РНК служит матрицей для синтеза комплементарной ДНК, которая может включаться в геном высших организмов (обратная транскрипция).
I. Структурная организация нуклеиновых кислот
В каждом живом организме присутствуют 2 типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). Молекулярная масса самой «маленькой» из известных нуклеиновых кислот — транспортной РНК (тРНК) составляет примерно 25 кД. ДНК — наиболее крупные полимерные молекулы; их молекулярная масса варьирует от 1 000 до 1 000 000 кД. ДНК и РНК состоят из мономерных единиц — нуклеотидов, поэтому нуклеиновые кислоты называют полинуклеотидами.
А. Строение нуклеотидов
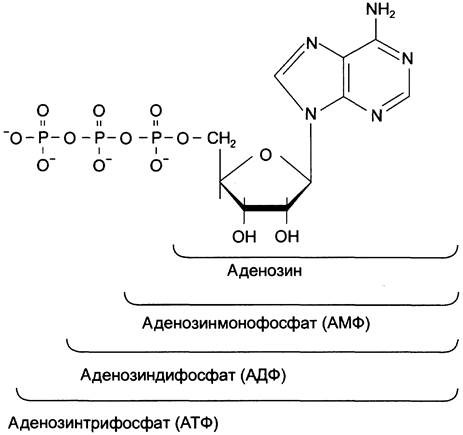
Каждый нуклеотид содержит 3 химически различных компонента: гетероциклическое азотистое основание, моносахарид (пентозу) и остаток фосфорной кислоты. В зависимости от числа имеющихся в молекуле остатков фосфорной кислоты различают нуклеозидмонофосфаты (НМФ), нуклеозиддифосфаты (НДФ), нуклеозидтрифосфаты (НТФ) (рис. 4-1).
Рис. 4-1. Нуклеозидмоно-, ди- и трифосфаты аденозина. Нуклеотиды — фосфорные эфиры нуклеозидов. Остаток фосфорной кислоты присоединён к 5'-углеродному атому пентозы (5'-фосфоэфирная связь).
В состав нуклеиновых кислот входят азотистые основания двух типов: пуриновые — аденин (А), гуанин (G) и пиримидиновые — цитозин (С), тимин (Т) и урацил (U).Нумерация атомов в основаниях записывается внутри цикла (рис. 4-2). Номенклатура нуклеотидов приведена в табл. 4-1.
Рис. 4-2. Пуриновые и пиримидиновые основания.
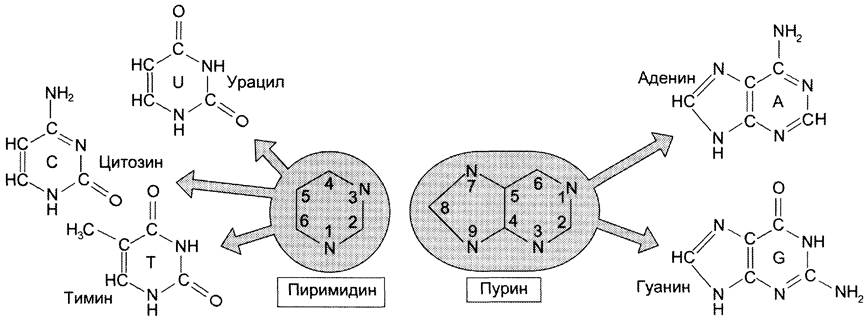
Таблица 4-1. Номенклатура нуклеотидов
Азотистое основание |
Нуклеозид |
Нуклеотид |
Трёхбуквенное обозначение |
Однобуквенный код |
Аденин |
Аденозин |
Аденозинмонофосфат |
АМФ |
А |
Гуанин |
Г уанозин |
Гуанозинмонофосфат |
ГМФ |
G |
Цитозин |
Цитидин |
Цитидинмонофосфат |
ЦМФ |
С |
Урацил |
Уридин |
Уридинмонофосфат |
УМФ |
U |
Тимин |
Тимидин |
Тимидинмонофосфат |
ТМФ |
Т |
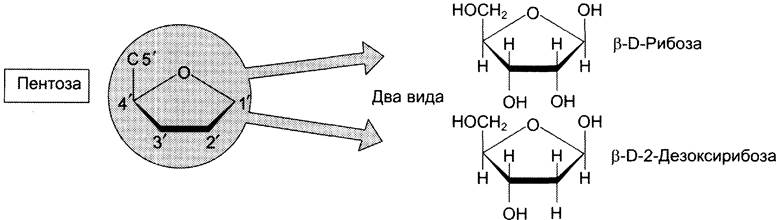
Пентозы в нуклеотидах представлены либо рибозой (в составе РНК), либо дезоксирибозой (в составе ДНК). Чтобы отличить номера атомов в пентозах от нумерации атомов в основаниях, запись производят с внешней стороны цикла и к цифре добавляют штрих (') — 1', 2', 3', 4' и 5' (рис. 4-3).
Рис. 4-3. Пентозы. Присутствуют 2 вида — β-D-рибоза в составе нуклеотидов РНК и β-D-2-дезоксирибоза в составе нуклеотидов ДНК.
Пентозу соединяет с основанием N-гликозидная связь, образованная С1-атомом пентозы (рибозы или дезоксирибозы) и N1-атомом пиримидина или N9-атомом пурина (рис. 4-4).
Рис. 4-4. Пуриновый и пиримидиновый нуклеотиды.

Нуклеотиды, в которых пентоза представлена рибозой, называют рибонуклеотидами, а нуклеиновые кислоты, построенные из рибонуклеотидов, — рибонуклеиновыми кислотами, или РНК. Нуклеиновые кислоты, в мономеры которых входит дезоксирибоза, называют дезоксирибонуклеиновыми кислотами, или ДНК. Нуклеиновые кислоты по своему строению относят к классу линейных полимеров. Остов нуклеиновой кислоты имеет одинаковое строение по всей длине молекулы и состоит из чередующихся групп — пентоза-фосфат-пентоза- (рис. 4-5). Вариабельными группами в полинуклеотидных цепях служат азотистые основания — пурины и пиримидины. В молекулы РНК входят аденин (А), урацил (U), гуанин (G) и цитозин (С), в ДНК — аденин (А), тимин (Т), гуанин (G) и цитозин (С). Уникальность структуры и функциональная индивидуальность молекул ДНК и РНК определяются их первичной структурой — последовательностью азотистых оснований в полинуклеотидной цепи.
Рис. 4-5. Фрагмент цепи ДНК.

Б. Структура дезоксирибонуклеиновой кислоты (ДНК)
Первичная структура ДНК — порядок чередования дезоксирибонуклеозидмонофосфатов (дНМФ) в полинуклеотидной цепи.
Каждая фосфатная группа в полинуклеотидной цепи, за исключением фосфорного остатка на 5'-конце молекулы, участвует в образовании двух эфирных связей с участием 3'- и 5'-углеродных атомов двух соседних дезоксирибоз, поэтому связь между мономерами обозначают 3', 5'-фосфодиэфирной.
Концевые нуклеотиды ДНК различают по структуре: на 5'-конце находится фосфатная группа, а на 3'-конце цепи — свободная ОН-группа. Эти концы называют 5'- и 3'-концами. Линейная последовательность дезоксирибонуклеотидов в полимерной цепи ДНК обычно сокращённо записывают с помощью однобуквенного кода, например -A-G-C-T-T-A-C-A- от 5'- к 3'-концу.
В каждом мономере нуклеиновой кислоты присутствует остаток фосфорной кислоты. При pH 7 фосфатная группа полностью ионизирована, поэтому in vivo нуклеиновые кислоты существуют в виде полианионов (имеют множественный отрицательный заряд). Остатки пентоз тоже проявляют гидрофильные свойства. Азотистые основания почти нерастворимы в воде, но некоторые атомы пуринового и пиримидинового циклов способны образовывать водородные связи.
Вторичная структура ДНК. В 1953 г. Дж. Уотсоном и Ф. Криком была предложена модель пространственной структуры ДНК. Согласно этой модели, молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная,полинуклеотидные цепи в ней антипараллельны (рис. 4-6), т. е. если одна из них ориентирована в направлении 3' —> 5', то вторая — в направлении 5' —> 3'. Поэтому на каждом из концов молекулы ДНК расположены 5'-конец одной цепи и 3'-конец другой цепи.
Рис. 4-6. Двойная спираль ДНК. Молекулы ДНК состоят из двух антипараллельных цепей с комплементарной последовательностью нуклеотидов. Цепи закручены относительно друг друга в правозакрученную спираль так, что на один виток приходится примерно 10 пар нуклеотидов.

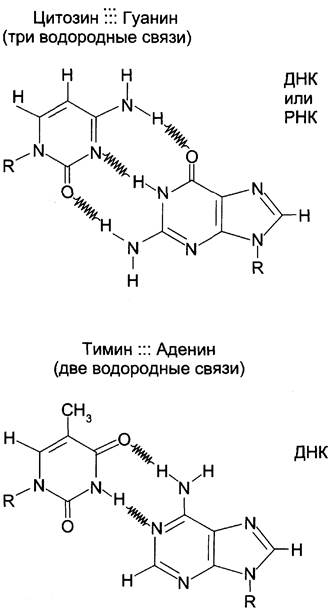
Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов — снаружи. Полинуклеотидные цепи удерживаются относительно друг друга за счёт водородных связей между комплементарными пуриновыми и пиримидиновыми азотистыми основаниями А и Т (две связи) и между G и С (три связи) (рис. 4-7). При таком сочетании каждая пара содержит по три кольца, поэтому общий размер этих пар оснований одинаков по всей длине молекулы. Водородные связи при других сочетаниях оснований в паре возможны, но они значительно слабее. Последовательность нуклеотидов одной цепи полностью комплементарна последовательности нуклеотидов второй цепи. Поэтому, согласно правилу Чаргаффа (Эрвин Чаргафф в 1951 г. установил закономерности в соотношении пуриновых и пиримидиновых оснований в молекуле ДНК), число пуриновых оснований (А + G) равно числу пиримидиновых оснований (Т + С).
Рис. 4-7. Пурин-пиримидиновые пары оснований в ДНК.
Комплементарые основания уложены в стопку в сердцевине спирали. Между основаниями двухцепочечной молекулы в стопке возникают гидрофобные взаимодействия,стабилизирующие двойную спираль.
Такая структура исключает контакт азотистых остатков с водой, но стопка оснований не может быть абсолютно вертикальной. Пары оснований слегка смещены относительно друг друга. В образованной структуре различают две бороздки — большую, шириной 2,2 нм, и малую, шириной 1,2 нм. Азотистые основания в области большой и малой бороздок взаимодействуют со специфическими белками, участвующими в организации структуры хроматина.
Третичная структура ДНК (суперспирализация ДНК)
Каждая молекула ДНК упакована в отдельную хромосому. В диплоидных клетках человека содержится 46 хромосом. Общая длина ДНК всех хромосом клетки составляет 1,74 м, но она упакована в ядре, диаметр которого в миллионы раз меньше. Чтобы расположить ДНК в ядре клетки, должна быть сформирована очень компактная структура. Компактизация и суперспирализация ДНК осуществляются с помощью разнообразных белков, взаимодействующих с определёнными последовательностями в структуре ДНК. Все связывающиеся с ДНК эукариотов белки можно разделить на 2 группы: гистоновые и негистоновые белки. Комплекс белков с ядерной ДНК клеток называют хроматином.
Гистоны — белки с молекулярной массой 11 — 21 кД, содержащие много остатков аргинина и лизина. Благодаря положительному заряду гистоны образуют ионные связи с отрицательно заряженными фосфатными группами, расположенными на внешней стороне двойной спирали ДНК.
Существует 5 типов гистонов. Четыре гистона Н2А, Н2В, Н3 и Н4 образуют октамерный белковый комплекс (Н2А, Н2В, Н3, Н4)2, который называют «нуклеосомный кор» (от англ. nucléosome соге). Молекула ДНК «накручивается» на поверхность гистонового октамера, совершая 1,75 оборота (около 146 пар нуклеотидов). Такой комплекс гистоновых белков с ДНК служит основной структурной единицей хроматина, её называют «нуклеосома». ДНК, связывающую нуклеосомные частицы, называют линкерной ДНК. В среднем линкерная ДНК составляет 60 пар нуклеотидных остатков. Молекулы гистона Н1 связываются с ДНК в межнуклеосомных участках (линкерных последовательностях) и защищают эти участки от действия нуклеаз (рис. 4-8).
Рис. 4-8. Структура нуклеосом. Восемь молекул гистонов (Н2А, Н2В, Н3, Н4)2 составляют ядро нуклеосомы, вокруг которого ДНК образует примерно 1,75 витка.
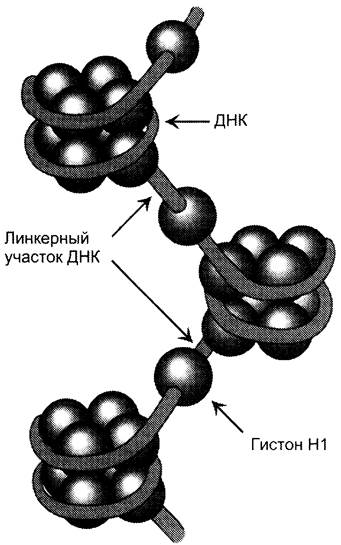
В ядре каждой клетки присутствует около 60 млн молекул каждого типа гистонов, а общая масса гистонов примерно равна содержанию ДНК. Аминокислотные остатки лизина, аргинина и концевые аминогруппы гистонов могут модифицироваться: ацетилироваться, фосфорилироваться, метилироваться или взаимодействовать с белком убиквитином (негистоновый белок). Модификации бывают обратимыми и необратимыми, они изменяют заряд и конформацию гистонов, а это влияет на взаимодействие гистонов между собой и с ДНК.
Активность ферментов, ответственных за модификации, регулируется и зависит от стадии клеточного цикла. Модификации делают возможными конформационные перестройки хроматина.
Негистоновые белки хроматина
В ядре эукариотической клетки присутствуют сотни самых разнообразных ДНК-связывающих негистоновых белков. Каждый белок комплементарен определённой последовательности нуклеотидов ДНК (сайт ДНК). К этой группе относят семейство сайт-специфических белков типа «цинковые пальцы» (см. раздел 1). Каждый «цинковый палец» узнаёт определённый сайт, состоящий из 5 нуклеотидных пар. Другое семейство сайт-специфических белков — гомодимеры. Фрагмент такого белка, контактирующий с ДНК, имеет структуру «спираль-поворот-спираль» (см. раздел 1). К группе структурных и регуляторных белков, которые постоянно ассоциированы с хроматином, относят белки высокой подвижности (HMG-белки — от англ. high mobility gel proteins). Они имеют молекулярную массу менее 30 кД и характеризуются высоким содержанием заряженных аминокислот. Благодаря небольшой молекулярной массе HMG-белки обладают высокой подвижностью при электрофорезе в полиакриламидном геле. К негистоновым белкам принадлежат также ферменты репликации, транскрипции и репарации. При участии структурных, регуляторных белков и ферментов, участвующих в синтезе ДНК и РНК, нить нуклеосом преобразуется в высоко конденсированный комплекс белков и ДНК. Образованная структура в 10 000 раз короче исходной молекулы ДНК.
В. Генетическая система митохондрий
Митохондрии — важнейшие органеллы клеток, осуществляющие синтез АТФ за счёт окисления субстратов. Митохондрии имеют собственный уникальный геном, наследуемый по материнской линии, так как он происходит из цитоплазмы яйцеклетки. Геном митохондрий сперматозоидов не попадает в оплодотворённую яйцеклетку.
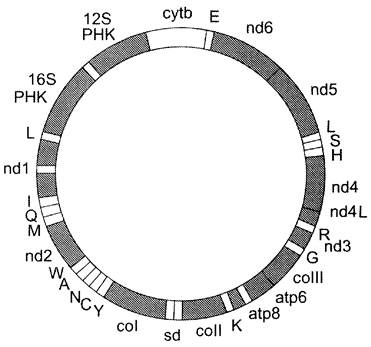
Митохондриальный геном человека представлен одной кольцевой молекулой ДНК из 16 569 нуклеотидных пар (рис. 4-9). Он кодирует 13 белков, используемых на построение структурно-функциональных компонентов митохондрий.
Рис. 4-9. Кольцевая молекула митохондриальной ДНК. Гены nd1-nd6, nd4I кодируют субъединицы NАОН-дегидрогеназного комплекса; ген соi-III — субъединицы цитохромоксидазы; ген суtb — цитохром b. Гены atp 8 и atp 6 кодируют субъединицы АТФ-синтазы (NАDН-дегидрогеназный комплекс, цитохро- моксидаза, цитохром b — белки, участвующие в энергетическом обмене). Остальные гены кодируют рибосомные (12S РНК и 16S РНК) и транспортные РНК соответствующих аминокислот, обозначенные латинскими буквами.
В митохондриях отсутствуют ферменты, ответственные за репарацию, поэтому митохондриальный геном содержит немало ошибок. Митохондрии эукариотов имеют очень маленькие рибосомы с константой седиментации 55S, тогда как рибосомы прокариотов — 70S.
Г. Структура рибонуклеиновых кислот (РНК)
Первичная структура РНК — порядок чередования рибонуклеозидмонофосфатов (НМФ) в полинуклеотидной цепи. В РНК, как и в ДНК, нуклеотиды связаны между собой 3',5'-фосфодиэфирными связями. Концы полинуклеотидных цепей РНК неодинаковы. На одном конце находится фосфорилированная ОН-группа 5'-углеродного атома, на другом конце — ОН-группа 3'-углеродного атома рибозы, поэтому концы называют 5'- и 3'-концами цепи РНК. Гидроксильная группа у 2'-углеродного атома рибозы делает молекулу РНК нестабильной. Так, в слабощелочной среде молекулы РНК гидролизуются даже при нормальной температуре, тогда как структура цепи ДНК не изменяется.
Вторичная структура РНК
Молекула рибонуклеиновой кислоты построена из одной полинуклеотидной цепи. Отдельные участки цепи РНК образуют спирализованные петли — «шпильки», за счёт водородных связей между комплементарными азотистыми основаниями А-U и G-С. Участки цепи РНК в таких спиральных структурах антипараллельны, но не всегда полностью комплементарны, в них встречаются неспаренные нуклеотидные остатки или даже одноцепочечные петли, не вписывающиеся в двойную спираль. Наличие спирализованных участков характерно для всех типов РНК.
Третичная структура РНК
Одноцепочечные РНК характеризуются компактной и упорядоченной третичной структурой, возникающей путём взаимодействия спирализованных элементов вторичной структуры. Так, возможно образование дополнительных водородных связей между нуклеотидными остатками, достаточно удалёнными друг от друга, или связей между ОН-группами остатков рибозы и основаниями. Третичная структура РНК стабилизирована ионами двухвалентных металлов, например, ионами Mg2+, связывающимися не только с фосфатными группами, но и с основаниями.
Основные типы РНК
В цитоплазме клеток присутствуют 3 типа рибонуклеиновых кислот — транспортные РНК (тРНК), матричные РНК (мРНК) и рибосомальные РНК (рРНК). Они различаются по первичной структуре, молекулярной массе, конформации, продолжительности жизни и, самое главное, по функциональной активности.
Транспортные РНК (тРНК)
Пространственную структуру любых тРНК, независимо от различий в последовательности нуклеотидов, описывают универсальной моделью «клеверного листа» (рис. 4-10). В каждой молекуле тРНК есть участки цепи, не участвующие в образовании водородных связей между нуклеотидными остатками. К ним, в частности, относят участок, ответственный за связывание с аминокислотой на 3'-конце молекулы и антикодон — специфический триплет нуклеотидов, взаимодействующий комплементарно с кодоном мРНК.
Рис. 4-10. Строение транспортных РНК. Спирализованные участки обозначены на рисунке пунктиром; «общие участки» одинаковы у всех тРНК; 1 — петля переменного размера; UН2 (дигидроурацил), Ψ (псевдоурацил) — минорные основания; антикодону всегда предшествует U (урацил), а после него всегда стоит минорное основание.

В состав нуклеотидов тРНК входят минорные основания (в среднем 10 — 12 оснований на молекулу). Они представлены метилированными основаниями, изомерами и аналогами пиримидинов (рис. 4-11).
Рис. 4-11. Минорные основания тРНК.

Минорные основания выполняют 2 функции: они делают тРНК устойчивыми к действию нуклеаз цитоплазмы и поддерживают определённую третичную структуру молекулы, так как не могут участвовать в образовании комплементарных пар, и препятствуют спирализации определённых участков в полинуклеотидной последовательности тРНК.
Матричные РНК (мРНК)
Первичная структура всех мРНК, независимо от уникальности их кодирующей последовательности, имеет одинаковое строение 5'- и 3'-концов. Так, на 5'- конце присутствует модифицированный нуклеотид 7-метилгуанозин- 5'-трифосфат (кэп). Несколько десятков нуклеотидов отделяют кэп от инициирующего кодона, обычно это триплет -AUG-. За кодирующим участком следует один из терминирующих кодонов — -UGA-, -UUA-, -UAG-. На 3'-конце большинства мРНК присутствует последовательность нуклеотидов из 100— 200 аденозинмонофосфатных остатков.
Рибосомальные РНК (рРНК)
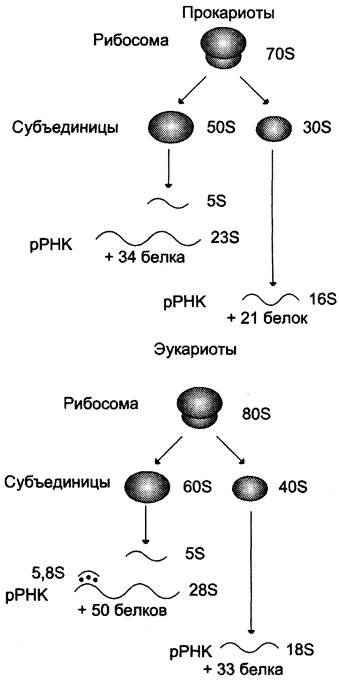
Рибосомальные РНК имеют многочисленные спирализованные участки. Различают рРНК — 5S, 5,8S, 28S и 18S (S — коэффициент седиментации). Рибосомальные РНК содержат несколько модифицированных нуклеотидов, чаще всего это метилированные производные азотистых оснований или рибозы (2'-метилрибоза). рРНК образуют комплексы с белками, которые называют рибосомами. Каждая рибосома состоит из двух субъединиц — малой (40S) и большой (60S). Субъединицы рибосом различаются не только набором рРНК, но и количеством и структурой белков (рис. 4-12).
Рис. 4-12. Строение эукариотических и прокариотических рибосом. Величина S характеризует скорость оседания частиц при ультрацентрифугировании и пропорциональна их молекулярной массе. Рибосома прокариотов (70S) состоит из 50S и 30S субъединиц, эукариотов (80S) — состоит из субъединиц 60S и 40S. Рибосомы эукариотов и прокариотов различаются по молекулярной массе субъединиц, количеству молекул рРНК, массе рРНК, количеству и разнообразию белков, способных связывать специфические лиганды.
Д. Гибридизация нуклеиновых кислот
Вторичная структура нуклеиновых кислот образуется за счёт слабых взаимодействий — водородных и гидрофобных. Поэтому если водный раствор ДНК нагреть до 100 °С, то связи, удерживающие две цепи двойной спирали вместе, разрушаются. В результате разрыва водородных и гидрофобных связей цепи ДНК расходятся. Этот процесс называют «денатурация». Однако если раствор, содержащий денатурированную ДНК, очень медленно охлаждать, то могут получиться двухспиральные структуры, идентичные исходным. Такой процесс получил название «ренативация».
На явлении денатурации и ренативации основан метод, называемый «молекулярная гибридизация». Процесс гибридизации может осуществляться между двумя любыми цепями нуклеиновых кислот (ДНК — ДНК, ДНК — РНК) при условии, что они содержат комплементарные последовательности нуклеотидов. Такие гибридные структуры можно выделить центрифугированием в градиенте плотности сахарозы или наблюдать в электронном микроскопе (рис. 4-13).
Рис. 4-13. Гибридизация нуклеиновых кислот. А - гибридизация ДНК-ДНК; Б - гибридизация ДНК-РНК.

Если раствор, содержащий образцы ДНК 1 и 2, выделенные из организмов разных видов, денатурировать, а затем провести ренативацию, то образуются двухспиральные структуры. Но наряду с исходными ДНК 1 и ДНК 2 образуются гибридные двойные спирали, содержащие цепь ДНК образца 1 и цепь ДНК образца 2, где присутствуют как спирализованные, так и неспирализованные участки. В неспирализованных участках фрагменты цепей ДНК не комплементарны, т. е. в ходе гибридизации получаются несовершенные гибриды. Методом молекулярной гибридизации можно установить:
✵ сходство и различие первичной структуры разных образцов нуклеиновых кислот;
✵ различие ДНК, выделенных из организмов разных видов;
✵ идентичность ДНК всех органов и тканей одного организма.
При проведении гибридизации ДНК-РНК были выделены гибридные молекулы, содержащие одну цепь ДНК и одну цепь РНК. Если для эксперимента были взяты ДНК и РНК (первичный транскрипт), выделенные из одного организма, то образовывались совершенные гибриды, потому что молекула РНК комплементарна цепи ДНК. Гибридизацией ДНК-РНК было впервые установлено, что все виды РНК клетки имеют на молекуле ДНК комплементарные участки.