БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ - Е. С. Северина - 2004
РАЗДЕЛ 7. ОБМЕН УГЛЕВОДОВ
III. Механизм трансмембранного переноса глюкозы и других моносахаридов в клетки
Моносахариды, образовавшиеся в результате переваривания, всасываются эпителиальными клетками тощей и подвздошной кишок с помощью специальных механизмов транспорта через мембраны этих клеток.
А. Всасывание моносахаридов в кишечнике
Транспорт моносахаридов в клетки слизистой оболочки кишечника может осуществляться разными способами: путём облегчённой диффузии и активного транспорта. В случае активного транспорта глюкоза и Nа+ проходят через мембраны с люминальной стороны, связываясь с разными участками белка-переносчика. При этом поступает в клетку по градиенту концентрации, и одновременно глюкоза транспортируется против градиента концентрации (вторично-активный транспорт, см. раздел 5). Следовательно, чем больше градиент Na+, тем больше поступление глюкозы в энтероциты. Если концентрация во внеклеточной жидкости уменьшается, транспорт глюкозы снижается. Градиент концентрации Na+, являющийся движущей силой активного симпорта, создаётся работой Na+, К+-АТФ-азы. Перенос в клетки слизистой оболочки кишечника по механизму вторично-активного транспорта характерен также для галактозы.
При разной концентрации глюкозы в просвете кишечника «работают» различные механизмы транспорта. Благодаря активному транспорту эпителиальные клетки кишечника могут поглощать глюкозу при её очень низкой концентрации в просвете кишечника. Если же концентрация глюкозы в просвете кишечника велика, то она может транспортироваться в клетку путём облегчённой диффузии. Таким же способом может всасываться и фруктоза. Следует отметить, что скорость всасывания глюкозы и галактозы гораздо выше, чем других моносахаридов. Способы транспорта моносахаридов через мембрану эпителиальных клеток кишечника представлены на рис. 7-18.
Рис. 7-18. Всасывание углеводов в кишечнике. Всасывание моносахаридов из кишечника происходит путём облегчённой диффузии с помощью специальных белков-переносчиков (транспортёров). Кроме того, глюкоза и галактоза транспортируются в энтероцит путём вторично-активного транспорта, зависимого от градиента концентрации ионов натрия. Белки-транспортёры, зависимые от градиента Na+, обеспечивают всасывание глюкозы из просвета кишечника в энтероцит против градиента концентрации. Концентрация Na+, необходимая для этого транспорта, обеспечивается Na+,К+-АТФ-азой, которая работает как насос, откачивая из клетки Na+ в обмен на К+. В отличие от глюкозы, фруктоза транспортируется системой, не зависящей от градиента натрия.
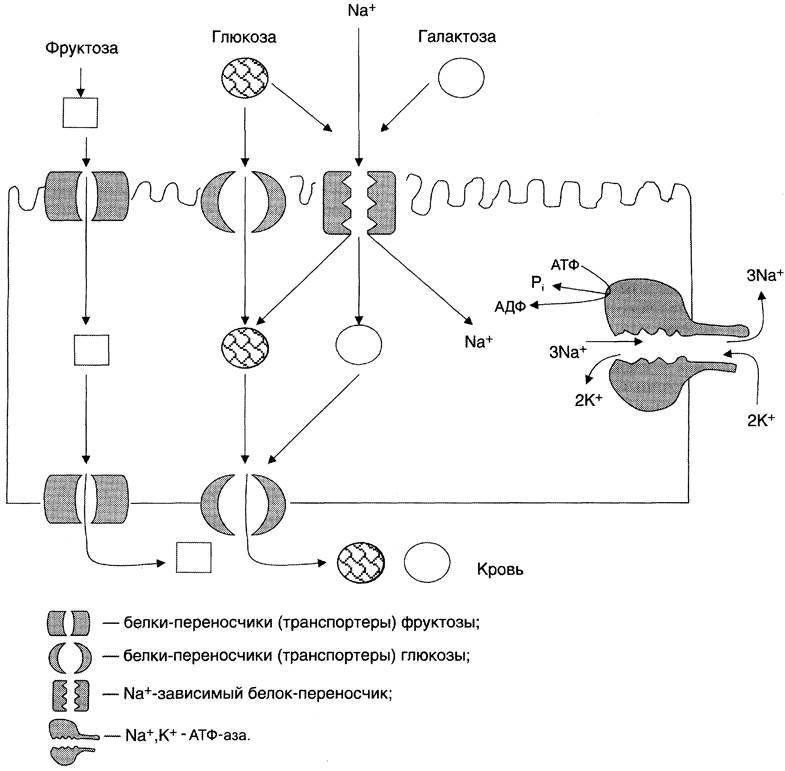
После всасывания моносахариды (главным образом, глюкоза) покидают клетки слизистой оболочки кишечника через мембрану, обращённую к кровеносному капилляру, с помощью облегчённой диффузии. Часть глюкозы (более половины) через капилляры кишечных ворсинок попадает в кровеносную систему и по воротной вене доставляется в печень. Остальное количество глюкозы поступает в клетки других тканей.
Б. Транспорт глюкозы из крови в клетки
Потребление глюкозы клетками из кровотока происходит также путём облегчённой диффузии. Следовательно, скорость трансмембранного потока глюкозы зависит только от градиента её концентрации. Исключение составляют клетки мышц и жировой ткани, где облегчённая диффузия регулируется инсулином (гормон поджелудочной железы). В отсутствие инсулина плазматическая мембрана этих клеток непроницаема для глюкозы, так как она не содержит белки- переносчики (транспортёры) глюкозы. Транспортёры глюкозы называют также рецепторами глюкозы. Например, описан транспортёр глюкозы, выделенный из эритроцитов. Это трансмембранный белок, полипептидная цепь которого построена из 492 аминокислотных остатков и имеет доменную структуру. Полярные домены белка расположены по разные стороны мембраны, гидрофобные располагаются в мембране, пересекая её несколько раз. Транспортёр имеет участок связывания глюкозы на внешней стороне мембраны. После присоединения глюкозы конформация белка изменяется, в результате чего глюкоза оказывается связанной с белком в участке, обращённом внутрь клетки. Затем глюкоза отделяется от транспортёра, переходя внутрь клетки (см. раздел 5).
Считают, что способ облегчённой диффузии по сравнению с активным транспортом предотвращает транспорт ионов вместе с глюкозой, если она транспортируется по градиенту концентрации.
Глюкозные транспортёры (ГЛЮТ) обнаружены во всех тканях. Существует несколько разновидностей ГЛЮТ (табл. 7-1), они пронумерованы в соответствии с порядком их обнаружения.
Таблица 7-1. Распределение белков-транспортёров глюкозы (ГЛЮТ)
Типы ГЛЮТ |
Локализация в органах |
ГЛЮТ-1 |
Преимущественно в мозге, плаценте, почках, толстом кишечнике |
ГЛЮТ-2 |
Преимущественно в печени, почках, β-клетках островков Лангерханса, энтероцитах |
ГЛЮТ-3 |
Во многих тканях, включая мозг, плаценту, почки |
ГЛЮТ-4 (инсулинзависимый) |
В мышцах (скелетной, сердечной), жировой ткани Содержится в отсутствие инсулина почти полностью в цитоплазме |
ГЛЮТ-5 |
В тонком кишечнике. Возможно, является переносчиком фруктозы. |
Структура белков семейства ГЛЮТ отличается от белков, транспортирующих глюкозу через мембрану в кишечнике и почках против градиента концентрации.
Описанные 5 типов ГЛЮТ имеют сходные первичную структуру и доменную организацию.
✵ ГЛЮТ-1 обеспечивает стабильный поток глюкозы в мозг;
✵ ГЛЮТ-2 обнаружен в клетках органов, выделяющих глюкозу в кровь. Именно при участии ГЛЮТ-2 глюкоза переходит в кровь из энтероцитов и печени. ГЛЮТ-2 участвует в транспорте глюкозы в β-клетки поджелудочной железы;
✵ ГЛЮТ-3 обладает большим, чем ГЛЮТ-1, сродством к глюкозе. Он также обеспечивает постоянный приток глюкозы к клеткам нервной и других тканей;
✵ ГЛЮТ-4 — главный переносчик глюкозы в клетки мышц и жировой ткани;
✵ ГЛЮТ-5 встречается, главным образом, в клетках тонкого кишечника. Его функции известны недостаточно.
Все типы ГЛЮТ могут находиться как в плазматической мембране, так и в цитозольных везикулах. ГЛЮТ-4 (и в меньшей мере ГЛЮТ-1) почти полностью находятся в цитоплазме клеток. Влияние инсулина на такие клетки приводит к перемещению везикул, содержащих ГЛЮТ, к плазматической мембране, слиянию с ней и встраиванию транспортёров в мембрану. После чего возможен облегчённый транспорт глюкозы в эти клетки. После снижения концентрации инсулина в крови транспортёры глюкозы снова перемещаются в цитоплазму, и поступление глюкозы в клетку прекращается (рис. 7-19).
Рис. 7-19. Влияние инсулина на перемещение транспортёров глюкозы из цитоплазмы в плазматическую мембрану. 1 — связывание инсулина с рецептором; 2 — участок инсулинового рецептора, обращённый внутрь клетки, стимулирует перемещение транспортёров глюкозы. 3, 4 — транспортёры в составе содержащих их везикул перемещаются к плазматической мембране клетки, включаются в её состав и переносят глюкозу в клетку.

Перемещение глюкозы из первичной мочи в клетки почечных канальцев происходит вторично-активным транспортом, подобно тому, как это осуществляется при всасывании глюкозы из просвета кишечника в энтероциты. Благодаря этому глюкоза может поступать в клетки даже в том случае, если её концентрация в первичной моче меньше, чем в клетках. При этом глюкоза реабсорбируется из первичной мочи почти полностью (99%).
Известны различные нарушения в работе транспортёров глюкозы. Наследственный дефект этих белков может лежать в основе инсулинонезависимого сахарного диабета (см. раздел 11). В то же время причиной нарушения работы транспортёра глюкозы может быть не только дефект самого белка. Нарушения функции ГЛЮТ-4 возможны на следующих этапах:
✵ передача сигнала инсулина о перемещении этого транспортёра к мембране;
✵ перемещение транспортёра в цитоплазме;
✵ включение в состав мембраны;
✵ отшнуровывание от мембраны и т. д.