Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ I. БІОМОЛЕКУЛИ ТА КЛІТИННІ СТРУКТУРИ
ГЛАВА 2. БІЛКИ І ПЕПТИДИ
2.2. БУДОВА Й АМІНОКИСЛОТНИЙ СКЛАД БІЛКІВ І ПЕПТИДІВ
Молекулярна маса білків. Білки є високомолекулярними сполуками; їх молекулярна маса (м.м.) коливається в межах від декількох тисяч до декількох мільйонів а.о.м. (дальтонів). Індивідуальні білки побудовані з декількох сотень амінокислотних залишків.
Білки можуть складатися з одного або декількох окремих поліпептидних ланцюгів, що об’єднані ковалентними (дисульфідними) та нековалентними зв’язками. Білки, в яких є один поліпептидний ланцюг, мають молекулярну масу від 5-6 до 50 кД; білки з більшою м.м. складаються, як правило, з декількох поліпептидних ланцюгів, що складають протомери (субодиниці) — мультиланцюгові (олігомерні) білки (табл. 2.1).
Таблиця 2.1. Молекулярна маса білків
Білок |
Молекулярна маса, кД |
Кількість субодиниць (протомерів) |
Інсулін |
5,7 |
1 |
Рибонуклеаза |
12,6 |
1 |
Пепсин |
35,5 |
1 |
Алкогольдегідрогеназа |
80,0 |
2 |
Піруваткіназа |
240,0 |
4 |
Фенілаланін-тРНК-синтетаза |
276,0 |
4 |
Пептиди (олігопептиди, поліпептиди) відрізняються від власне білків молекулярною масою (меншою 5-6 кД) та відповідними фізико-хімічними властивостями.
Форма білкових молекул
Поліпептидні ланцюги, що лежать в основі ковалентної структури білкових молекул, здатні до формування впорядкованих конформацій, які стабілізуються водневими та іншими слабкими фізико-хімічними зв’язками. Ці високовпорядковані конформації створюють певні рівні структурної організації білків (див. нижче), що відображуються в різних формах будови білкових молекул.
За формою молекул білки поділяються на глобулярні — кулеподібні та фібрилярні — з витягнутою формою молекули.
Детальніше питання утворення глобулярних та фібрилярних білків будуть розглянуті при обговоренні механізмів формування вищих рівнів структурної організації білків.
Амінокислотний склад білків і пептидів
При гідролізі природних білків та пептидів вивільнюється близько 20 різних α-L-амінокислот, розміщення кожної з яких у поліпептидному ланцюгу кодується триплетом нуклеотидів у ДНК геному.
Амінокислоти, що входять до складу природних білків та пептидів (протеїногенні амінокислоти), мають загальну хімічну структуру, яка представлена наведеною звичайною структурною (а) та проекційною (б) формулами (рис. 2.1).
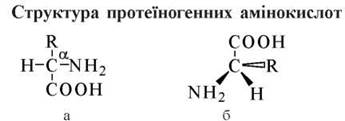
Рис. 2.1. Структурна (а) та проекційна (б) формули протеїногенних амінокислот. Наведено проекційну формулу L-амінокислоти.
Структурні особливості протеїногенних амінокислот:
1) аміногрупа, іон водню та боковий ланцюг (R-група) зв’язані з атомом вуглецю, що міститься в α-положенні відносно карбоксильної групи, тобто природні амінокислоти є α-амінокислотами; деякі амінокислоти (лізин, аргінін) мають додаткову аміногрупу, що розташована в кінцевому положенні (ω-) радикала R;
2) за своєю абсолютною конфігурацією протеїногенні амінокислоти є стереоізомерами L-ряду (L-амінокислотами). D-амінокислоти до складу природних білків не входять; вони зустрічаються в бактеріальних та рослинних об’єктах, входять до складу деяких антибіотиків (граміцидин, актиноміцин D). Оптичні ізомери амінокислот диференціюються за смаком (L — гіркі або без смаку, D — солодкі), що свідчить про стереоспецифічність смакових рецепторів.
Класифікація протеїногенних амінокислот
Природні α-амінокислоти можуть поділятися на класи залежно від хімічної будови бічного радикалу R:
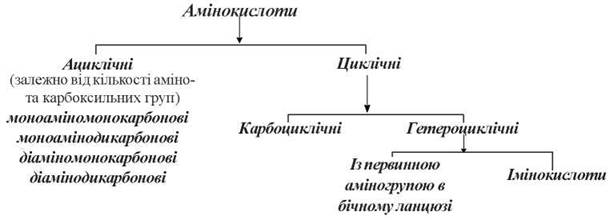
Сучасна раціональна класифікація, що базується на полярності та заряді радикалу R, передбачає чотири класи амінокислот (табл. 2.2):
I — амінокислоти з неполярними (гідрофобними) R-групами;
II — амінокислоти з полярними (гідрофільними) незарядженими R-групами;
III — амінокислоти з негативно зарядженими R-групами (кислі амінокислоти);
IV — амінокислоти з позитивно зарядженими R-групами (основні амінокислоти).
Крім зазначених у таблиці 2.2 двадцяти амінокислот, у складі деяких білків виявлено похідні цих амінокислот, зокрема 4-гідроксипролін, 5-гідроксилізин, N-метилізин, 3-метилгістидин, фосфосерин, фосфотреонін, дийодтирозин. Хімічна модифікація (гідроксилювання, фосфорилювання, йодування) відповідних амінокислот відбувається вже після їх включення в поліпептидні ланцюги (посттрансляційна модифікація білків).
Таблиця 2.2. Амінокислоти, що входять до складу білків
Назва |
Міжнародний символ |
Структурна формула |
pІ |
Амінокислоти з неполярними R-групами |
|||
Аланін |
Ala (A) |
|
6,02 |
Валін |
Val (V) |
|
5,97 |
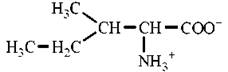
Лейцин |
Leu (L) |
|
5,98 |
Ізолейцин |
Ile (I) |
|
6,02 |
Метіонін |
Met (M) |
|
5,75 |
Пролін |
Pro (P) |
|
6,10 |
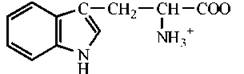
Триптофан |
Tip (W) |
|
5,88 |
Фенілаланін |
Phe (F) |
|
5,98 |
Амінокислоти з полярними незарядженими R-групами |
|||
Гліцин |
Gly (G) |
|
5,97 |
Серин |
Ser (S) |
|
5,68 |
Треонін |
Thr (T) |
|
6,53 |
Цистеїн |
Cys (C) |
|
5,02 |
Тирозин |
Tyr (Y) |
|
5,65 |
Аспарагін |
Asn (N) |
|
5,41 |
Глутамін |
Gln (Q) |
|
5,65 |
Амінокислоти з негативно зарядженими R-групами |
|||
Аспарагінова кислота |
Asp (D) |
|
2,97 |
Глутамінова кислота |
Glu (E) |
|
3,22 |
Амінокислоти з позитивно зарядженими R-групами |
|||
Лізин |
Lys (K) |
|
9,74 |
Аргінін |
Arg (R) |
|
10,76 |
Гістидин |
His (H) |
|
7,58 |
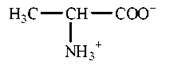
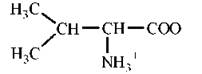
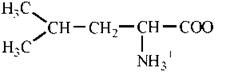
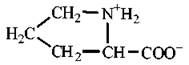
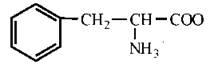
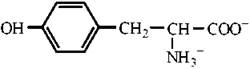
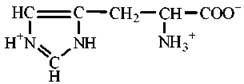
Властивості протеїногенних амінокислот
1. Кислотно-основні властивості амінокислот.
Амінокислоти є амфотерними електролітами, що можуть дисоціювати з утворенням іонних форм — аніона або катіона. У водному середовищі амінокислоти існують у вигляді рівноважної суміші, що складається з аніонної, катіонної форм та біполярного іона (цвіттер-іона) — рис. 2.2.
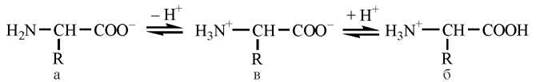
Рис. 2.2. Аніонна (а), катіонна (б) та біполярна (в) форми амінокислот у водних розчинах.
Зазначені реакції утворення аніонів, катіонів та біполярних іонів амінокислот повністю відповідають тільки схемі кислотно-основної дисоціації моноаміномоно-карбонових амінокислот, що мають по одній а-амінній та α-карбоксильній групі. У цьому найпростішому випадку рівновага між позитивно та негативно зарядженими молекулами може теоретично досягатися вже в нейтральних розчинах, тобто при рН=7.
Разом із тим, деякі амінокислоти мають бокові ланцюги R, що містять додаткові функціональні групи, здатні до дисоціації:
- кислотні групи Asp, Glu;
- основні групи Lys, Arg, His.
Таким чином, сумарний заряд молекул амінокислот (та, відповідно, білків і пептидів, до складу яких вони входять) визначається взаємовідношенням між кількістю вільних кислотних та основних груп, ступенем їх дисоціації (рКа) та рН середовища.
У кислих розчинах переважає катіонна форма амінокислот (молекули заряджені позитивно), в лужних розчинах — аніонна (амінокислоти заряджені негативно). Ці фізико-хімічні властивості амінокислот визначають їх здатність до електрофорезу — розділенню у високовольтному постійному електричному полі. При рівновазі позитивних та негативних зарядів молекула амінокислоти перебуває в ізоелектричному стані. Характерне для кожної амінокислоти значення рН, при якому амінокислота має сумарний нульовий заряд, називається рН ізоелектричної точки (рІ).
2. Полярність молекул амінокислот.
Залежно від полярності бічних радикалів R (табл. 2.2), амінокислоти в більшій або меншій мірі взаємодіють із диполями води, тобто проявляють гідрофільні або гідрофобні властивості.
Полярність функціональних груп амінокислот разом з їх кислотно-основними властивостями визначають особливості структури, більшість фізико-хімічних та, відповідно, біологічних властивостей білків, що синтезуються з цих амінокислот.
3. Оптичні властивості амінокислот.
α-Атом вуглецю всіх протеїногенних амінокислот, за винятком гліцину, зв’язаний із чотирма різними функціональними групами (асиметричний атом) і є хіральним центром молекули; на основі цього, протеїногенні амінокислоти є оптично активними сполуками, тобто здатні до обертання площини поляризованого світла.
4. Здатність до утворення кислото-амідних зв’язків.
Характерною хімічною особливістю амінокислот є здатність їх α-амінної та α-карбоксильної груп утворювати кислото-амідний (пептидний) зв’язок за рахунок відщеплення елементів молекули води, тобто вступати до реакції поліконденсації:
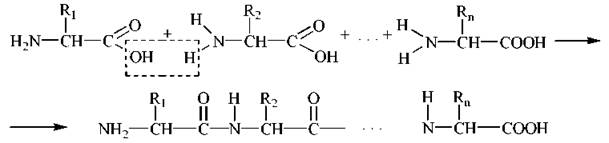
Поліаміди, що утворюються в зазначених реакціях, отримали назву пептидів (дипептидів, трипептидів ... олігопептидів ... поліпептидів, відповідно).
Структура пептидної групи
Чотири атоми, що входять до складу пептидної групи (-СО-NH-), розміщені в одній геометричній площині, тобто є компланарними. Кисень карбонільної групи та водень NH-групи розташовані в трансположенні (рис. 2.3).
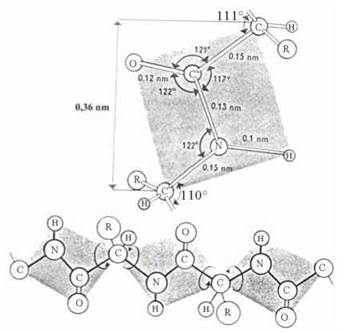
Рис. 2.3. Компланарне розміщення атомів пептидної групи в молекулах пептидів та білків.
Довжина зв’язку між атомами вуглецю карбонільної групи та азоту амідної групи дорівнює 0,132 нм, тобто цей зв’язок коротший звичайного одинарного зв’язку C-N (0,147 нм) і є приблизно на 50 % подвійним. Такий характер подвійного зв’язку зумовлений спряженням вільної пари p-електронів азоту з п-електронами подвійного зв’язку С=О (р,п-спряження) та утворенням резонансної структури:
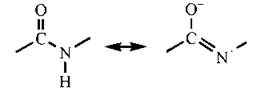
Виходячи з будови пептидного зв’язку та пептидної групи, вільне обертання в пептидному ланцюзі можливе тільки навколо груп -CHR, що розташовані між окремими компланарними пептидними групами (рис. 2.3). Ці структурні обмеження, разом із здатністю певних функціональних груп пептидних ланцюгів до сильних та слабких взаємодій, детермінують особливості утворення упорядкованих конформацій молекул білків, що розглянуті нижче.
5. Хімічні реакції, що використовуються для аналізу амінокислот.
Завдяки різноманітності своїх функціональних груп молекули α-амінокислот можуть вступати в хімічні реакції, які застосовуються в аналітичній та клінічній біохімії для ідентифікації і кількісного визначення окремих амінокислот. Ці реакції (так звані «кольорові реакції») використовуються для визначення як вільних амінокислот, що містяться як у біологічних об’єктах (плазмі крові, сечі тощо), так і в межах аналізу амінокислотного складу білків та пептидів.
Нінгідринова реакція
Нінгідрин (трикетогідринденгідрат) при нагріванні з а-амінокислотами спричиняє їх декарбоксилювання з утворенням NH3, СО2 та альдегіду — продукту окислювального декарбоксилювання амінокислоти. У подальшому аміак, що вивільнився, реагує з відновленим нінгідрином, утворюючи комплекс синьо-фіолетового кольору з максимумом поглинання при λmax = 570 нм. За допомогою нінгідринової реакції можливо детектувати 1 нмоль амінокислоти:
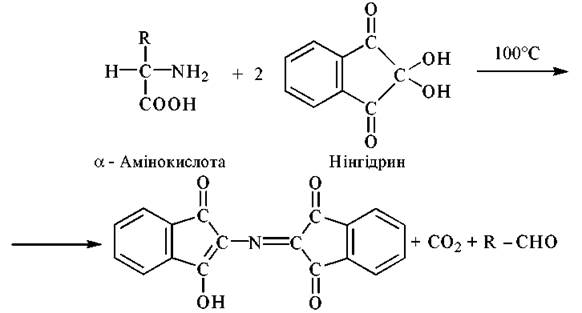
Флуорескамінова реакція
Високочутливим реагентом на α-амінокислоти є також флуорескамін, який утворює з амінокислотами флуоресцуючі комплекси. Флуорескамінова реакція є більш чутливою, ніж нінгідринова, і дозволяє визначати амінокислоти в кількостях 10-50 пмолей.
Спектрофотометричне або спектрофлуоро-метричне вимірювання комплексів амінокислот із нінгідрином або флуорескаміном дозволяє кількісно визначати амінокислоти не тільки як вільні метаболіти, а й у складі білкових гідролізатів після їх хроматографічного розділення, що використовується в аналізі первинної структури білків та пептидів.
Крім зазначених, у клінічній біохімії застосовують такі «кольорові реакції» амінокислот:
- ксантопротеїнова реакція — характерна для бензольного ядра циклічних амінокислот (фенілаланіну, тирозину, триптофану), яке нітрується при дії концентрованої азотної кислоти з утворенням нітросполук жовтого кольору;
- реакція Мілона — специфічна реакція на тирозин (амінокислоту, що містить фенольний гідроксил). В умовах нагрівання фенолів та їх похідних із реактивом Мілона (суміш нітратів ртуті (I) та (II)) утворюються ртутні похідні цегляно-червоного кольору;
- реакція Сакагучі — реакція, що застосовується для ідентифікації гуанідинової групи аргініну. При взаємодії гуанідину з α-нафтолом та гіпохлоритом натрію в лужних умовах утворюються сполуки з червоним забарвленням;
- реакція Ерліха — застосовується для виявлення індольного кільця триптофану, яке при реакції з n-диметиламінобензальдегідом у кислому середовищі дає сполуки з фіолетовим забарвленням;
- реакція Фоля — реакція, характерна для сірковмісних амінокислот. При кіп’ятінні розчину білка або відповідних амінокислот із лугом у присутності плюмбіту натрію утворюється чорно-бурий осад сульфіду свинцю.