Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ V. ГОРМОНИ В СИСТЕМІ МІЖКЛІТИННОЇ ІНТЕГРАЦІЇ ФУНКЦІЙ ОРГАНІЗМУ
ГЛАВА 24. ГОРМОНАЛЬНА РЕГУЛЯЦІЯ МЕТАБОЛІЗМУ ТА БІОЛОГІЧНИХ ФУНКЦІЙ КЛІТИНИ. II. ГОРМОНИ - ПОХІДНІ ПЕПТИДІВ ТА АМІНОКИСЛОТ
Білково-пептидні гормони — численна група фізіологічно активних сполук із гормональними та нейромедіаторними властивостями, що продукуються в різних органах і тканинах та за своєю хімічною природою є поліпептидами (простими білками, глікопротеїнами, низькомолекулярними пептидами).
Основні класи білково-пептидних гормонів, що синтезуються в ендокринній системі: гормони гіпофіза, гіпоталамуса, підшлункової залози. Близькими за механізмами дії до білково-пептидних гормонів є біорегулятори — похідні амінокислот (біогенні аміни) — катехоламіни, серотонін, гістамін, мелатонін.
Простими пептидами є також біорегулятори, що належать до гістогормонів:
- опіоїдні пептиди головного мозку;
- пептиди гастроінтестинальної системи;
- компоненти кінінової та ренін-ангіотензинової систем;
- гормони та медіатори імунної системи і функціонально близькі до них пептидні фактори росту, цитомедини;
- натрійуретичні пептиди серця, мозку тощо.
24.1. ГІПОТАЛАМО-ГІПОФІЗАРНА СИСТЕМА
Центральною ендокринною залозою організму людини та вищих тварин є гіпофіз — орган, що контролює ендокринну діяльність більшості підпорядкованих їй (периферійних) залоз внутрішньої секреції. В свою чергу, функціональна активність гіпофіза регулюється нейроендокринними клітинами спеціальних ядер гіпоталамуса. Гормони та/або нейромедіатори (чи модуляторні нейропептиди) гіпоталамічних та інших підкіркових ядер головного мозку контролюють секрецію (а в деяких випадках — біосинтез, продукування) гіпофізарних гормонів.
А. Гормони гіпоталамуса
Гіпоталамус є зоною головного мозку, яка регулює активність гіпофіза і периферійних ендокринних залоз шляхом продукції в нейросекреторних клітинах специфічних гіпоталамічних, гіпофізотропних гормонів та дії нейротрансмітерів, що контролюють функції підпорядкованих залоз внутрішньої секреції через симпатичну та парасимпатчну нервову систему.
Згідно з рекомендаціями Комісії з біохімічної номенклатури Міжнародного біохімічного союзу (1974), гормони гіпоталамуса, що сприяють вивільненню (release — англ.) певних гормонів гіпофіза, позначають як «ліберини » (рилізинг-гормони), а біорегулятори, які гальмують вивільнення гіпофізарних гормонів — «статини» (інгібіруючі гормони).
На даний час встановлені утворення та секреція в гіпоталамусі таких пептидних гіпофізотропних гормонів:
Соматоліберин (соматотропін-рилізинг-гормон, СТГ-РГ) — під впливом СТГ- РГ стимулюється продукція і вивільнення в гіпофізі гормону росту.
Соматостатин (СС; гормон, що інгібірує виділення гормону росту) — гормон, під впливом якого гальмуються продукція і вивільнення гормону росту; крім гіпоталамуса, соматостатин синтезується також в острівках підшлункової залози та інших клітинних утвореннях шлунково-кишкового тракту, де виконує специфічні фізіологічні функції.
Пролактостатин (пролактин-інгібіруючий гормон, ПІГ) — пептид, що гальмує продукцію пролактину та має гонадоліберинову активність.
Тироліберин (тиротропін-рилізинг-гормон; ТРГ) — гормон, під впливом якого стимулюється продукція і вивільнення тиреотропного гормону гіпофіза.
Гонадоліберин (гонадотропін-рилізинг-гормон; ГнРГ; люліберин) — під впливом ГнРГ стимулюється синтез і вивільнення гонадотропних гормонів ФСГ та ЛГ.
Кортиколіберин (кортикотропін-рилізинг-гормон) — гормон, що стимулює вивільнення кортикотропіну.
За хімічною природою ліберини та статини є поліпептидами, що секретуються певними нейронами гіпоталамуса, та, надходячи разом із біогенними амінами (дофаміном, серотоніном, норадреналіном) і іншими нейромедіаторами через систему портального кровообігу або нейрональні аксони в аденогіпофіз, регулюють його специфічні гормональні функції. Продукція гіпоталамічних гормонів є об’єктом складної регуляції з боку центральної нервової системи, нейромедіатори якої (біогенні аміни, нейропептиди, медіаторні амінокислоти) здійснюють модулюючий вплив на функціональну активність ядер гіпоталамуса.
Б. Гормони передньої частки гіпофіза
Передня частка гіпофіза (аденогіпофіз) продукує значну кількість гормонів, які стимулюють фізіологічні та біохімічні процеси в різних тканинах-мішенях, в тому числі активують дію інших ендокринних залоз (тропна функція гіпофізарних гормонів). Виходячи з особливостей молекулярної генетики, біосинтезу та структурно- функціональних властивостей, гормони аденогіпофіза утворюють три групи:
I група — «гормон росту-пролактин-хоріонічний соматомамотропін»;
II група — глікопротеїни — «тропні гормони гіпофіза»;
III група — похідні «проопіомеланокортину».
I. Група гормону росту — є біохімічним угрупованням («сімейством») білкових гормонів, які мають значну гомологію первинної структури (амінокислотних послідовностей) та близькість функціонально-біохімічних ефектів. Це гормони, що складаються у більшості тваринних видів із 190-199 амінокислотних залишків і мають ростостимулюючу та лактогенну активності.
1. Гормон росту (соматотропін, соматотропний гормон, СТГ) — простий білок, що складається з одного поліпептидного ланцюга (191 амінокислотний залишок; м.м. — 21,5 кД) і має два внутрішньомолекулярні дисульфідні зв’язки. Гормон синтезується в соматотрофних клітинах, що складають близько 50 % клітин аденогіпофіза. Ген, який контролює експресію СТГ, знаходиться в 17-й хромосомі каріотипу людини. СТГ синтезується у вигляді декількох прогормонів, що відповідають за наявність певної молекулярної гетерогенності гормону росту.
Біологічні властивості СТГ
Головна функція СТГ — стимуляція постнатального росту організму; ця складна біологічна функція реалізується за рахунок різноманітного спектра впливу гормону на біосинтез білка, вуглеводний та ліпідний метаболізм:
а) вплив на біосинтез білка — характеризується анаболічною спрямованістю: СТГ стимулює транспорт амінокислот у клітини та процеси транскрипції і трансляції в гормон-чутливих тканинах (переважно в м’язах, хрящах, кістках, печінці, сполучній тканині тощо); в цілому вплив СТГ на організм призводить до позитивного азотистого балансу, тобто переважання процесів синтезу над катаболізмом білків та амінокислот;
б) вплив на обмін вуглеводів та ліпідів — визначається «контраінсулярними ефектами» гормону: введення СТГ супроводжується гіперглікемією (гіперглюкоземією), яка є результатом як зменшення утилізації глюкози клітинами (гальмування транспорту глюкози з екстрацелюлярного простору та інгібірування її гліколітичного окислення), так і активації її продукції в ході глюконеогенезу; в адипоцитах жирової тканини СТГ активує реакції ліполізу, що призводить до стимуляції виходу НЕЖК та гліцеролу в плазму крові.
Лактогенні властивості СТГ пов’язані з його здатністю до взаємодії з лактогенними рецепторами молочної залози; в здоровому організмі ці ефекти СТГ не мають суттєвого значення і проявляють себе в умовах патології гіпофіза.
Соматомедини
Особливістю біологічної дії СТГ є її опосередкованість через синтез у печінці та біологічні ефекти двох поліпептидних факторів росту (соматомединів) — ІФР-1 та ІФР-2 («інсуліноподібних факторів росту 1 та 2» — пептидів, що складаються з 70 та 67 амінокислотних залишків, відповідно).
Первинна структура соматомединів нагадує будову молекули проінсуліну. Ген, що кодує синтез ІФР-1, локалізується в 12-й хромосомі людини, ген ІФР-2 — в 11-й хромосомі, в безпосередній близькості до гена, який кодує інсулін. Обидва фактори росту стимулюють реплікацію ДНК, що проявляється підсиленням включення в ДНК тимідину, транскрипцію та трансляцію в тканинах-мішенях СТГ.
Стимуляція росту тканин під впливом соматомединів виражена в 50-100 разів сильніше, порівняно з інсуліном, при цьому ростостимулюючі властивості СТГ в найбільшій мірі корелюють з ефектами ІФР-1: особини з дефіцитом цього соматомедину позбавлені здатності до нормального росту. Ефекти ІФР реалізуються через рецептори, локалізовані на плазматичних мембранах клітин-мішеней — рецептори 1-го типу, що мають тирозин-кіназну активність та рецептори 2-го типу, активація яких призводить до включення ефекторних систем клітини через G-білки.
Регуляція секреції СТГ
Продукція та секреція гормону росту знаходяться під позитивним та негативним нейрогуморальним контролем: виділення СТГ з гіпофіза стимулюється соматоліберином та гальмується соматостатином. Крім того, активуючий вплив на виділення СТГ гіпофізом справляють такі фізіологічно активні сполуки, як дофамін, серотонін, вазопресин, естрогени, агоністи γ-адренергічних рецепторів та у-аміномасляної кислоти.
Вивільнення СТГ (подібно до інших гормонів гіпоталамо-гіпофізарної системи) відбувається в пульсуючому режимі — максимум синтезу гормону спостерігається через 60-90 хв від початку сну (ранні стадії фази глибокого, «повільнохвилевого» сну); процес сну вважається фізіологічним стимулятором секреції соматотропіну («Діти ростуть під час сну»!).
Патологія, пов’язана з гормоном росту.
акромегалія — захворювання, розвиток якого спричиняється збільшеною продукцією гормону росту у дорослих осіб; захворювання характеризується патологічно диспропорційним збільшенням кісток скелета (особливо кінцівок, щелеп та інших компонентів черепа), м’яких тканин, внутрішніх органів; у жінок молодого віку захворювання проявляє себе галактореєю, що спричинена збільшеною продукцією пролактину або власною лактогенною активністю СТГ. Етіологія захворювання пов’язана з наявністю пухлини — аденоми гіпофіза (соматотропіноми);
гігантизм — прояв надмірної секреції гормону росту в дитячому та підлітковому віці, що призводить до збільшеного росту людини (умовно — вище 190 см); пролонгування в дорослому віці аномально високої продукції СТГ призводить у таких пацієнтів до розвитку акромегалії;
карликовість (нанізм, дварфізм) — затримка росту (у чоловіків — нижче 130 см, у жінок — нижче 120 см), яка спричиняється гетерогенними факторами, пов’язаними як із зменшенням синтезу СТГ та, відповідно, ІФР-1 («карлики з дефіцитом СТГ»), так і з порушеннями реактивності тканин на дію гормону: «карлики Ларона» — особини з відсутністю рецепторів СТГ в печінці та пігмеї, у яких має місце молекулярна патологія в пострецепторній трансдукції гормонального сигналу.
2. Пролактин (лактогенний гормон, мамотропін, лютеотропний гормон) — простий білок , що складається з одного поліпептидного ланцюга (м.м. 23 кД). Гормон продукується в ацидофільних клітинах аденогіпофіза — лактотрофах, кількість і розміри яких збільшуться під час вагітності.
Пролактин бере участь в ініціації та стимуляції лактації у жінок, причому дія гормону проявляється лише на тлі сенсибілізації клітин молочної залози жіночими статевими гормонами.
Пухлини, що складаються з пролактинсинтезуючих клітин гіпофіза, призводять у жінок до аменореї та галактореї, у чоловіків — до деяких видів безплідності.
Синтез та секреція пролактину гальмуються дофаміном та специфічним інгібіруючим нейропептидом гіпоталамуса — пролактостатином, що має також властивості гонадоліберину (гонадоліберинасоційований пептид, ГАП).
3. Хоріонічний соматомамотропін (ХС; плацентарний лактоген) — гормон, що проявляє лактогенну та лютеотропну активності, а за метаболічними ефектами близький до соматотропіну. Істинна фізіологічна роль ХС в організмі людини не з’ясована.
II. Група тропних гормонів гіпофіза — включає в себе сполуки глікопротеїнової природи: тиреотропний та гонадотропний гормони гіпофіза і плаценти (фолікулостимулювальний гормон, лютеїнізуючий гормон, хоріонічний гонадотропін).
Кожен із гормонів групи, що розглядається, є димером і складається з двох субодиниць — α та β, які сполучені між собою нековалентними зв’язками. Молекулярна маса цих гормонів—близько 30 кД, вуглеводні (олігосахаридні) групи складають 15-30 % маси молекули; до складу олігосахаридних радикалів входять залишки галактози, манози, фукози, галактозаміну, глюкозаміну, сіалових кислот.
У всіх чотирьох глікопротеїнових гормонів α-субодиниці однакові, а β-субодиниці різні і визначають біологічну специфічність гормону. α-Субодиниці є коротшими (містять у собі 89-96 залишків амінокислот) і сполучені з 2 вуглеводними радикалами, а β-субодиниці — довші і варіабельні за складом (містять 113-119 амінокислотних залишків і декілька вуглеводних радикалів):
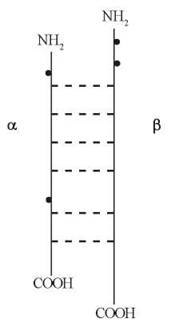
Схема будови глікопротеїнових гормонів (жирними точками позначені вуглеводні залишки).
Гени, що кодують α- і β-субодиниці глікопротеїнових гормонів, розміщуються на різних хромосомах — 6-й та 1-й, відповідно.
Тиреотропний гормон (ТТГ; тиротропін) — подібно до інших глікопротеїнових гормонів гіпофіза є димером типу αβ (м.м. близько 30 кД). Гормон синтезується базофільними клітинами передньої частки — тиротрофами, які складають 3-5 % клітинного складу аденогіпофіза.
Основна біологічна функція ТТГ — підтримка функціональної активності (синтезу тиреоїдних гормонів) та структури щитовидної залози. Ефекти ТТГ щодо тиреоцитів реалізуються за мембранним механізмом: взаємодія гормону з мембранними рецепторами через посередництво різних G-білків призводить до активації аденілатциклази та фосфоліпази С. Таким чином, активація ТТГ гормонопродукуючої функції щитовидної залози досягається через декілька вторинних месенджерів: цАМФ, інозитол-1,4,5-трифосфату та діацилгліцеролу.
Вивільнення гіпофізом ТТГ позитивно модулюється специфічним гіпоталамічним гормоном тироліберином. З іншого боку, гормони щитовидної залози тироксин та трийодтиронін гальмують тиреотропну функцію гіпофіза шляхом гальмування секреції гіпоталамусом тироліберина.
Патологія продукції ТТГ можлива за умов виникнення аденом гіпофіза, що секретують тиретропін, і проявляється симптомами тиреотоксикозу (див. нижче).
Гонадотропні гормони
Гонадотропіни є гормонами, що забезпечують нормальний гаметогенез та продукцію відповідних статевих гормонів у чоловічому та жіночому організмах. Це глікопротеїни, які за молекулярною будовою є також димерами типу αβ. Синтез гонадотропінів відбувається в базофільних клітинах гіпофіза — гонадотрофах, які складають 10-15 % від загальної кількості клітин аденогіпофіза. В період вагітності відбувається фізіологічна гіперплазія гонадотрофів.
1. Фолікулостимулюючий гормон (ФСГ; фолітропін) — білок з м.м. 33 кД. Мішенями для ФСГ є фолікулярні клітини яєчників та клітини Сертолі сім’яників. Передача хімічного сигналу при дії ФСГ здійснюється за рахунок активації аденілатциклази.
2. Лютеїнізуючий гормон (ЛГ; лютропін; гормон, що стимулює інтерстиціальні клітини Лейдига — ГСІК) — білок з м.м. 29 кД. Рецептори для ЛГ локалізовані на плазматичних мембранах клітин яєчників (у жіночому організмі) та клітин Лейдига сім’яників (у чоловічому організмі). Як і в разі ФСГ, вторинним месенджером в дії ЛГ на ефекторні системи клітин є цАМФ.
3. Хоріонічний гонадотропін (ХГ) — білок з м.м.близько 37 кД; синтезується трофобластом плаценти.
Біологічна роль гонадотропінів — полягає в регуляції функцій статевої сфери людини як в препубертатному, пубертатному періодах, так і у дорослих особин, процесів як гаметогенезу, так і продукції статевих гормонів.
У жіночому організмі — ФСГ та ЛГ контролюють функціонування менструального циклу (див. главу 25), стимулюючи ріст фолікулів, синтез у них естрогенів (переважна дія ФСГ), овуляцію та утворення жовтого тіла (дія ФСГ на тлі присутності ЛГ), персистенцію жовтого тіла і продукцію прогестерону (дія ЛГ та/або ХГ — в умовах вагітності).
У чоловічому організмі — ФСГ сприяє активації процесів сперматогенезу: спричиняє проліферацію клітин Сертолі та сперматогенного епітелію, підвищує чутливість клітин Лейдигу до дії ЛГ (за рахунок збільшення в них кількості ЛГ-чутливих рецепторів). Мішенню для дії ЛГ (ГСІК) є інтерстиціальні клітини Лейдига, в яких гормон стимулює біосинтез із холестерину тестостерону — основного статевого гормону чоловічого організму.
Регуляція секреції гонадотропних гормонів здійснюється гіпоталамусом за рахунок виділення гонадоліберину, що стимулює синтез та секрецію аденогіпофізом як ФСГ, так і ЛГ. Властивості гонадоліберину має також пролактостатин (гонадоліберинасоційований пептид, ГАП). Подібно до інших тропних гормонів аденогіпофіза, продукція ФСГ та ЛГ контролюється за принципом негативного зворотного зв’язку концентраціями статевих гормонів: збільшення рівнів естрогенів та андрогенів у крові гальмує виділення ЛГ, тоді як секреція ФСГ пригнічується прогестероном. Зменшення функціональної активності статевих залоз (менопауза, хірургічна або променева кастрація) супроводжується стимуляцією секреції гонадотропінів.
Порушення синтезу, секреції та/або рецепції гонадотропінів спостерігаються при численних спадкових і набутих захворюваннях, що можуть призводити до різних клінічних форм ураження репродуктивної функції організму людини.
III. Група проопіомеланокортину — гормони, що входять до цієї групи (сімейства) є продуктами посттрансляційного процесингу біологічного попередника — прогормону проопіомеланокортину(ПОМК).
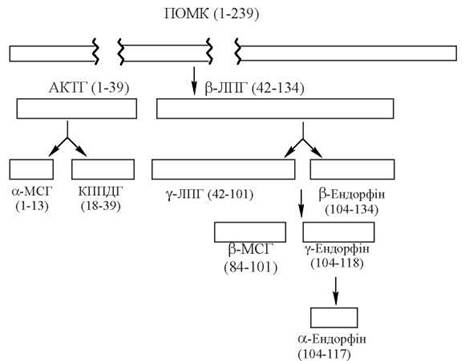
Утворення фізіологічно активних пептидів із проопіомеланокортину.
ПОМК синтезується в базофільних клітинах гіпофіза; це глікопротеїн, що складається з 239 амінокислотних залишків (м.м. близько 30 кД) і є попередником багатьох фізіологічно активних пептидів гормональної та нейромедіаторної дії, які утворюються з ПОМК за рахунок обмеженого протеолізу та реакцій ковалентної модифікації (глікозилювання, ацетилювання).
Процесинг ПОМК відбувається в передній та проміжній частках гіпофіза (проміжна частка активна лише в ембріональний період та у жінок на пізніх стадіях вагітності) і деяких периферійних тканинах: плаценті, кишечнику, чоловічому статевому тракті. Основними продуктами цього процесингу є:
- адренокортикотропний гормон (АКТГ), що в свою чергу є попередником меланоцитостимулюючого гормону (α-МСГ) та кортикотропіноподібного пептиду проміжної частки гіпофіза (КППДГ);
- β-ліпотропний гормон (β-ЛПГ), що є попередником γ-ліпотропіну (γ-ЛПГ), β-МСГ та ендорфінів (β, γ, α).
1. Адренокортикотропний гормон (АКТГ; кортикотропін) — одноланцюговий пептид, що складається з 39 амінокислотних залишків (м.м. 4,5 кД).
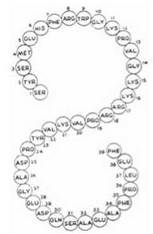
Первинна структура АКТГ людини.
Головними мішенями АКТГ є клітини кори наднирникових залоз, відносно яких гормон проявляє два типи біологічної активності: стимуляцію стероїдогенезу та підтримання маси наднирникових залоз.
(1) Вплив АКТГ на біосинтез наднирникових гормонів зумовлений активацією цАМФ-залежних протеїнкіназ, що каталізують ключові реакції перетворення холестерину на прегненолон — попередник С21-стероїдів кортикостероїдів. Основним ефектом такої дії АКТГ є стимуляція синтезу глюкокортикоїдів (головним чином, кортизолу), проте в умовах тривалої стимуляції кори наднирникових залоз кортикотропіном (хвороба Іценко-Кушинга, введення фармакологічних препаратів АКТГ) відбувається також активація синтезу мінералокортикоїдів та андрогенів.
(2) Збільшення маси кори наднирникових залоз при дії АКТГ (переважно сітчастої та пучкової зон) здійснюється також через стимуляцію цАМФ-залежних протеїнкіназ, які активують шляхом фосфорилювання певні рибосомальні білки та стимулюють синтез ДНК та РНК, необхідних для утворення нових клітин.
Позанаднирниковозалозні ефекти АКТГ полягають у стимуляції ліполізу в жировій тканині, активації поглинання амінокислот та глюкози м’язами. АКТГ має також певну меланоцитостимулюючу дію (збільшення пігментації шкіри), яка зумовлена наявністю в його первинній структурі тетрапептидного угруповання, загального з МСГ (див. нижче).
Вивільнення гіпофізом АКТГ є об’єктом складного нейрогуморального контролю, в якому беруть участь як прямі, так і зворотні регуляторні зв’язки: позитивними регуляторами є гормон гіпоталамуса кортиколіберин, а також вазопресин, адреналін, ангіотензин II; негативним регулятором — глюкокортикоїд кортизол.
2. Ліпотропний гормон (ЛПГ; ліпотропін) — група пептидів, що мають властивості активувати ліполіз в адипоцитах жирової тканини і мобілізацію жирних кислот.
У гіпофізі людини знайдені β- та γ-ліпотропіни; оскільки специфічна ліполітична активність цих пептидів незначна у порівнянні з дією інших гормонів та біорегуляторів, існує думка, що їх фізіологічне значення полягає в утворенні ендорфінів.
3. Ендорфіни — представники групи опіоїдних нейропептидів (α-, β-, γ- та δ-ендорфіни), що виконують функції нейромедіаторів, ендогенних знеболювальних факторів та модуляторів певних важливих психофізіологічних процесів у пептидергічних структурах головного мозку (глава 33).
4. Меланоцитостимулюючий гормон (МСГ) — група пептидів (α-, β-, γ-МСГ), які продукуються в проміжній частці гіпофіза і стимулюють функціональну активність меланоцитів шкіри, збільшуючи її пігментацію. Біохімічні механізми дії МСГ полягають у підвищенні активності тирозинази — ферменту, що бере участь у перетворенні тирозину на пігменти меланіни. Ця специфічна біологічна активність різних МСГ зумовлена наявністю в первинній структурі гормону характерного тетрапептиду - His-Phe-Arg-Trp-, що присутній також у молекулі АКТГ.
У гіпофізах ссавців присутні різні типи МСГ; фізіологічна функція гормону в людини недостатньо з’ясована у зв’язку з редукцією проміжної частки гіпофіза у дорослої людини — α-МСГ виявляється у людини лише в гіпофізах плода, при пухлинах гіпофіза та іноді в період вагітності.
В. Гормони задньої частки гіпофіза
Гормонами задньої частки гіпофіза є вазопресин та окситоцин. Це нейрогіпофізарні гормони, оскільки задня частка гіпофіза є лише місцем їх накопичення, а біосинтез відбувається в супраоптичному та паравентрикулярному ядрах гіпоталамуса. Гормони, що синтезовані в гіпоталамічних нейросекреторних клітинах, транспортуються в задню частку гіпофіза по аксонах нейрогіпофізарного тракту; транспортною формою гормонів є їх гранули в комплексі з білком нейрофізином.
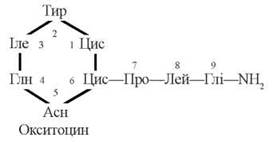
Вазопресин та окситоцин є циклічними пептидами, що складаються з 9 амінокислотних залишків (нонапептиди); первинна структура двох гормонів розрізняється лише залишками амінокислот, які містяться в 3-му та 8-му положеннях.
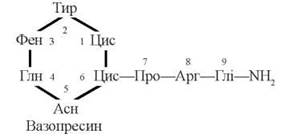
(1) Вазопресин
Біологічні функції вазопресину (антидіуретичного гормону, АДГ) пов’язані з регуляцією осмолярності та осмотичного тиску рідин організму.
Біохімічна основа фізіологічних ефектів вазопресину полягає в стимуляції реабсорбції води в дистальних канальцях нирок. Крім того, гормон сприяє підтриманню артеріального тиску за рахунок прямого впливу на судинну стінку, підсилює гліюгеноліз у печінці та м’язах, спричиняє агрегацію тромбоцитів та виділення ними факторів коагуляції.
Молекулярні механізми дії вазопресину ґрунтуються на наявності двох типів рецепторів цього активного пептиду:
V1-рецепторів (локалізовані на мембранах гепатоцитів, гладеньких м’язів судин, тромбоцитів), що спряжені з фосфоліпазою С, активація якої включає фосфоінозитидний цикл (утворення ІФ3, ДАГ) та збільшує концентрацію в клітинах іонів Са2+;
V2-рецептори (локалізовані на мембранах епітеліальних клітин трубочок та петель Генле нефронів), що спряжені з активацією аденілатциклази та утворенням цАМФ; послідовна дія цього біохімічного каскаду призводить до збільшення проникності мембран клітин-мішеней для води.
Порушення синтезу, транспортування та вивільнення в гіпоталамусі або зниження чутливості рецепторів нефронів до вазопресину призводять до розвитку важкого захворювання — нецукрового діабету, клінічними проявами якого є виділення значної кількості сечі (декількох літрів на добу) з низькою щільністю та постійне відчуття спраги.
(2) Окситоцин
Фізіологічна дія окситоцину полягає в активації скорочення м’язів матки (стимуляції пологової діяльності) та скорочення міоепітеліальних клітин, що оточують альвеоли молочної залози (забезпеченні надходження молока з альвеол у вивідні протоки під час лактації). Ефекти гормону на чутливі клітини реалізуються через аденілатциклазну систему за обов’язковою участю іонів Са2+.