Биохимия и молекулярная биология - Белясова Н.А. 2002
Молекулярные основы и механизмы наследственности
Экспрессия генов
Транскрипция ДНК
Наследственная информация, записанная языком последовательности нуклеотидов, у большинства организмов (кроме РНК-содержащих вирусов) хранится в ДНК. Последовательность триплетов нуклеотидов в генах определяет последовательность аминокислот в полипептидах или рибонуклеотидов в молекулах транспортных и рибосомальных РНК. Для того чтобы генетическая программа реализовалась (синтезировались нужные белки и РНК), требуется участие аппарата экспрессии генов. Под процессом экспрессии генов понимают синтез матричных РНК и белков. При этом мРНК выступают в роли посредников между ДНК и белком: синтез белка всегда осуществляется на однонитевых мРНК (при участии рибосом), а сами мРНК всегда синтезируются на двухнитевых ДНК. Оба процесса относятся к числу матричных и подлежат регуляции, которая существенным образом сказывается на уровне клеточного метаболизма.
Экспрессия всех генов начинается с транскрипции их нуклеотидной последовательности. Транскрипция — это процесс перевода информации, записанной на языке последовательности дезоксирибонуклеотидов в смысловой цепи ДНК на язык последовательности рибонуклеотидов в мРНК. При этом определенный участок одной из двух цепей ДНК (антисмысловой) используется как матрица для синтеза РНК путем комплементарного спаривания оснований.
Ферментами, катализирующими процесс транскрипции, служат ДНК-зависимые РНК-полимеразы. Причем у прокариот, например, в клетках кишечной палочки обнаружен лишь один тип этого фермента, который синтезирует все три типа РНК (мРНК, тРНК, рРНК). В отличие от них эукариоты имеют три разные ДНК-зависимые РНК-полимеразы, каждая из которых ответственна за транскрипцию генов, кодирующих разные типы клеточных РНК. Наилучшим образом процесс транскрипции, а также его ферментативное оснащение изучены у прокариот. Бактериальные РНК-полимеразы — это сложные белки, состоящие из нескольких разных субъединиц. Наиболее изученный фермент — холофермент РНК-полимераза E. coli, который содержит пять разных полипептидных субъединиц: две a-цепи, одну ß- и одну ß'-цепи, σ- и ω-цепи. Альтернативная форма фермента, называемая кором или миниферментом, лишена σ-субъединицы. Кор-фермент катализирует большинство реакций транскрипции ДНК в РНК, однако не может инициировать синтез РНК в нужном месте, поскольку не способен узнавать промоторные сайты. Точное связывание и инициация в промоторах происходят только после добавления к корферменту σδ-субъединицы и образования холофермента.
Как и другие матричные процессы, транскрипция включает 3 стадии: инициацию, элонгацию и терминацию.
Инициация транскрипции. Для этого процесса необходимы: холофермент, специальная последовательность нуклеотидов в ДНК (промотор) и набор нуклеозидтрифосфатов. Транскрипция инициируется при образовании стабильного комплекса между холоферментом и специфической последовательностью, называемой промотором и располагающейся в начале всех транскрипционных единиц. Промотор — это участок молекулы ДНК, состоящий примерно из 40 пар нуклеотидов и расположенный непосредственно перед участком инициации транскрипции. В нем различают две важные и достаточно консервативные по составу последовательности. Одна из них состоит из шести или семи нуклеотидов (чаще ТАТААТ) и расположена на расстоянии примерно 10 нуклеотидов от первого транскрибируемого нуклеотида (+1); этот сигнал обычно обозначают как -10-последовательность, или Прибнов-Бокс — в честь ее первооткрывателя. В данном сайте РНК-полимераза связывается с ДНК. Вторая последовательность расположена на расстоянии ~ 35 нуклеотидов до сайта инициации и служит участком распознавания промотора РНК-полимеразой (рис. 3.1).
Когда РНК-полимераза связывается с промотором, происходит локальное расплетение двойной спирали ДНК и образуется открытый промоторный комплекс. В нем происходит копирование последовательности нуклеотидов смысловой, или (+)-цепи ДНК, имеющей направление 5' → 3'. При этом синтез мРНК всегда начинается с нуклеотидов А или G. Вторая, антисмысловая цепь ДНК, служит матрицей для синтеза цепочки РНК (рис. 3.2).
Транскрипция аналогична репликации в том смысле, что порядок присоединения рибонуклеотидов определяется комплементарным спариванием оснований (рис. 3.2). После формирования первых нескольких фосфодиэфирных связей (обычно 5—10) δ-субъединица отделяется от инициирующего комплекса, и дальнейшая транскрипция осуществляется с помощью корфермента.
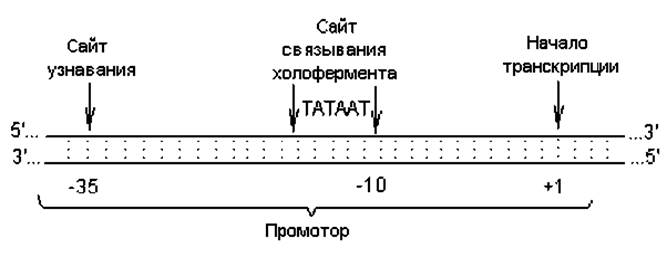
Рис. 3.1. Структура промотора E. coli (объяснения в тексте)
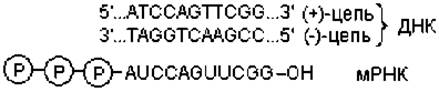
Рис. 3.2. Матричный принцип процесса транскрипции
Элонгация транскрипции. Растущая цепь РНК остается связанной с ферментом и спаренной своим растущим концом с участком матричной цепи. Остальная часть образовавшейся цепи не связана ни с ферментом, ни с ДНК. По мере продолжения транскрипции движущийся вдоль цепи ДНК корфермент действует подобно застежке «молния», расплетая двойную спираль, которая замыкается позади фермента, и восстанавливается ее исходная дуплексная структура. «Раскрытая» ферментом область ДНК простирается всего на несколько пар нуклеотидов (рис. 3.3).
Наращивание РНК идет в направлении от 5'- к 3'-концу вдоль матричной (-) цепи, ориентированной в направлении 3' → 5', т. е. антипараллельно. Транскрипция непрерывно продолжается до тех пор, пока фермент не достигнет сайта терминации транскрипции.
Терминация транскрипции. Последовательности ДНК, являющиеся сигналами остановки транскрипции, называются транскрипционными терминаторами. Они содержат инвертированные повторы, благодаря чему 3'-концы РНК-транскриптов складываются с образованием шпилек разной длины (рис. 3.4).
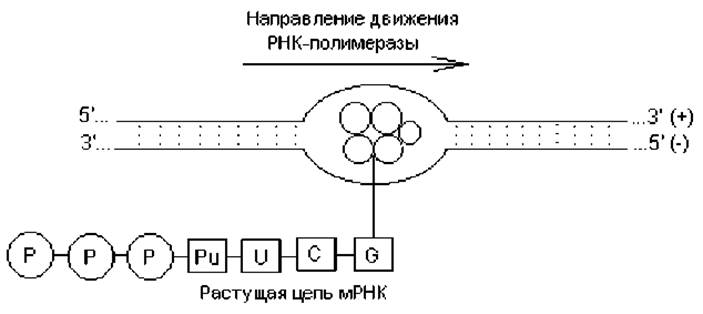
Рис. 3.3. Транскрипция ДНК: в локальном участке «расплетенной» ДНК работает корфермент, синтезируя цепь мРНК в направлении 5'Р → 3'ОН
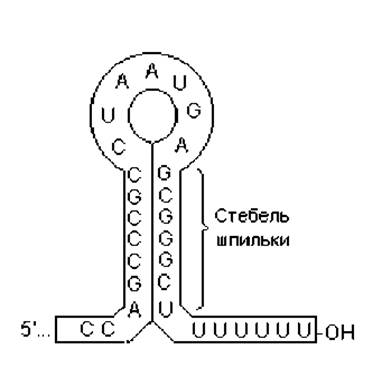
Рис. 3.4. Пример шпильки в составе мРНК, на которой заканчивается процесс транскрипции. В составе стебля шпильки присутствуют комплементарные нуклеотиды
Обнаружены два типа сигналов терминации — p-зависимый и р-независимый терминаторы. р — это олигомерный белок, прочно связывающийся с РНК и в этом состоянии гидролизующий АТР до ADP и неорганического фосфата. В одной из моделей действие р-белка объясняется тем, что он связывается с синтезируемой цепью РНК и перемещается вдоль нее в направлении 5' → 3' к месту синтеза РНК; необходимая для его перемещения энергия выделяется при гидролизе АТР. Если р-белок наталкивается на образующуюся в РНК шпильку, он останавливает полимеразу, которая могла бы продолжить транскрипцию. Механизм p-независимой терминации изучен хуже, в нем остается много неясного.
В большинстве случаев первичные транскрипты, образующиеся описанным выше способом, не являются зрелыми молекулами РНК, а требуют процесса созревания, который называется процессингом РНК. Процессинг сильно отличается для прокариотических и эукариотических РНК.
У прокариот первичные транскрипты, сформированные при транскрипции генов, кодирующих белки, функционируют в качестве мРНК без последующей модификации или процессинга. Причем трансляция мРНК часто начинается даже до завершения синтеза 3'-конца транскрипта. Совсем иная ситуация наблюдается для молекул прокариотических рРНК и тРНК. В этом случае кластеры рРНК- или тРНК-генов часто транскрибируются с образованием единой цепи РНК. Для формирования зрелых функциональных форм должны произойти специфическое надрезание первичных РНК-транскриптов и модификация. Эти молекулярные события и называют процессингом РНК или посттранскрипционной модификацией. Начальное расщепление первичных транскриптов на фрагменты, содержащие либо тРНК, либо 16S-, 23S- или 5S-рРНК-последовательности, осуществляет эндонуклеаза РНК-аза Ш. Ее мишенями служат короткие дуплексы РНК, образующиеся при внутримолекулярном спаривании оснований в последовательностях, фланкирующих каждый из РНК-сегментов. Эти комплементарные последовательности формируют шпильки, в составе которых РНК-аза вносит разрывы, после чего лишние последовательности спейсерных областей удаляются другими РНК-азами. Молекулы тРНК вначале синтезируются в виде про-тРНК, которая на ~ 20 % длиннее, чем зрелая. Лишние последовательности, расположенные у 5' и 3'-концов, удаляются рибонуклеазами Q и P. Кроме этого, для образования зрелой функциональной тРНК, по-видимому, должны произойти специфическая модификация оснований и присоединение одного, двух или всех трех нуклеотидов 3'-ССА-конца (акцепторная ветвь).
Созревание РНК у эукариот осуществляется гораздо сложнее. Во-первых, у эукариот существует ядро, которое отделено от цитоплазмы ядерной мембраной. В ядре осуществляется образование первичных транскриптов, которые имеют бульшую длину, чем цитоплазматическая мРНК, участвующая в трансляции. Следовательно, образованию зрелой мРНК у эукариот должно предшествовать удаление интронов из последовательности гяРНК-транскрипта (этот процесс называется сплайсингом от англ. to splice — сплетать, сращивать). После удаления последовательностей, соответствующих интронам, происходит соединение участков, которые транскрибированы с экзонов. Сплайсинг катализируется комплексами белков с РНК (мяРНП), которые, взаимодействуя с гяРНК, образуют сплайсому. Полагают, что каталитической активностью в сплайсоме обладает РНК-составляющая. Такие РНК называют рибозимами. Место сплайсинга определяется в сплайсомах с высокой точностью, поскольку ошибка даже в 1 нуклеотид может привести к искажению структуры белка. Для точного узнавания в составе интронов есть специфические последовательности — сигналы.
Кроме сплайсинга, мРНК у эукариот подвергается модификации: на 5'-конце синтезируется «кэп» (шапочка) — структура, представляющая собой метилированный остаток гуанозинтрифосфата, который защищает РНК от гидролиза 5'-экзонуклеазами. На 3'-конце про-мРНК синтезируется полиаденилатная последовательность длиной 150—200 нуклеотидов, которая называется «шлейф». Эти структуры принимают участие в регуляции экспрессии эукариотических генов. Процессинг рРНК и тРНК у эукариот осуществляется аналогично таковому у прокариот.