Биохимия и молекулярная биология - Белясова Н.А. 2002
Структура и функции клеточных компонентов
Биомембраны
Транспорт веществ через мембраны
Жизнедеятельность клетки связана с постоянным обменом ее содержимого с окружающей средой. Точно так же и внутри клетки происходит перемещение веществ между органеллами или компартментами. Все эти события связаны с преодолением основного барьера для веществ — мембраны, ограничивающей органеллу или саму клетку. При этом следует помнить, что главная функция биомембран — избирательность транспорта для различных веществ и ионов. Возможные способы транспорта через мембраны можно разделить на 4 основных типа: пассивная диффузия, облегченная диффузия, активный транспорт и цитозы.
Пассивная диффузия. Это процесс транспорта через мембраны веществ из области с большей их концентрацией в область с меньшей концентрацией (по химическому градиенту), в котором не принимают участия транспортные белки и не затрачивается энергия. С помощью такого способа через мембрану транспортируются малые незаряженные молекулы, например молекулы газов, некоторые анестезирующие вещества, а также вода. Чтобы пересечь бислой, молекула должна преодолеть поверхностное натяжение на границе мембраны, проникнуть в бислой, продиффундировать через него и выйти с противоположной стороны, вновь преодолев энергетический барьер на границе раздела фаз. Этим и объясняется избирательная проницаемость липидного бислоя для небольших молекул неэлектролитов. Удивительным является факт весьма легкого и быстрого проникновения воды через мембраны: показано, что молекуле воды требуется для пересечения бислоя всего 1 мкс. Для объяснения этого феномена в последнее время появляются основанные на некоторых экспериментальных данных предположения о том, что в мембранах все же существуют какие-то белковые проводящие пути для воды, либо молекулы воды пользуются локальными дефектами в структуре бислоев.
Перемещение одних только молекул воды через полупроницаемую мембрану можно рассматривать как частный вид диффузии — осмос. Под осмосом понимают переход молекул воды из области с высоким водным потенциалом и низкой концентрацией растворенного вещества в область с низким водным потенциалом и высокой концентрацией растворенного вещества (рис. 4.9). В этом случае молекулы воды будут переходить из гипотонического раствора в гипертонический до тех пор, пока не наступит равновесие и оба раствора не станут изотоническими по отношению друг к другу.
Чтобы обозначить величину уменьшения водного потенциала, вызванного присутствием растворенных веществ, используют термин «осмотическое давление». Под осмотическим давлением понимают давление, которое следует приложить к раствору, чтобы остановить осмотическое поступление воды в него через полупроницаемую мембрану. Повышение концентрации растворенного вещества увеличивает осмотическое давление и уменьшает водный потенциал раствора.
Перемещение воды через плазматические мембраны клеток в соответствии с законами осмоса создает организмам немалые проблемы, особенно для водных обитателей. Поэтому осморегуляция (поддержание водного потенциала в клетке на постоянном уровне) является важной стороной функциональной деятельности большинства организмов, и на ее осуществление зачастую тратится значительная доля запасенной клеткой энергии.
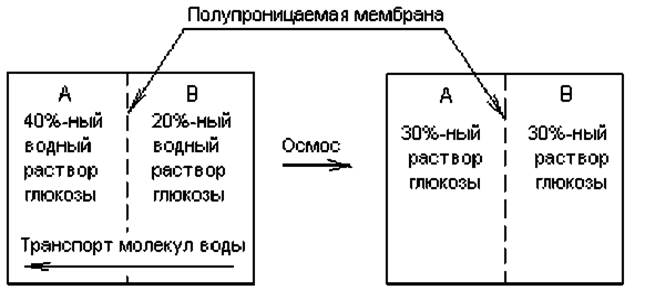
Рис 4.9. Движение молекул воды в соответствии с закономерностями осмоса. Раствор А содержит более высокую концентрацию глюкозы, чем раствор В, и является по отношению к нему гипертоническим. Соответственно раствор В по отношению к раствору А является гипотоническим. Результатом осмотического перемещения воды становится уравнивание концентраций и воды, и глюкозы: растворы становятся изотоническими по отношению друг к другу
Скорость диффузии веществ через мембраны зависит от многих причин: растворимости вещества в мембране, коэффициента диффузии в мембране, а также разности концентрации вещества снаружи и внутри клетки (градиента концентрации) (рис. 4.10).
Облегченная диффузия. Этот вид транспорта осуществляется с помощью транспортных белков по электрохимическому градиенту (разность электрических потенциалов и концентраций веществ) без затрат энергии. Это селективный перенос веществ — вещество будет транспортировано через мембрану лишь в том случае, если для него в мембране имеется функционирующий транспортный белок. Поскольку в облегченной диффузии задействованы белки, этот процесс, в отличие от пассивной диффузии, может достигать эффекта насыщения. Стадия насыщения (рис. 4.10) характеризует состояние, когда все транспортные белки для данного вещества насыщены субстратом и скорость транспорта этого вещества достигает максимума.
С помощью облегченной диффузии через мембрану транспортируются многие вещества, в том числе гидрофильные молекулы: углеводы, аминокислоты, нуклеотиды, различные ионы и др. При этом скорость транспорта значительно превышает скорость пассивной диффузии (рис. 4.10). Принципиально возможны два пути переноса веществ и ионов через мембрану: с помощью переносчиков и каналов. Поскольку трансмембранное перемещение белков в биомембранах не обнаружено, предложена модель, описывающая работу переносчиков — механизм «пинг-понг». Согласно этому механизму, транспорт веществ связан с конформационными изменениями в структуре белка-переносчика, которые индуцируются связыванием транспортируемого вещества (рис. 4.11).
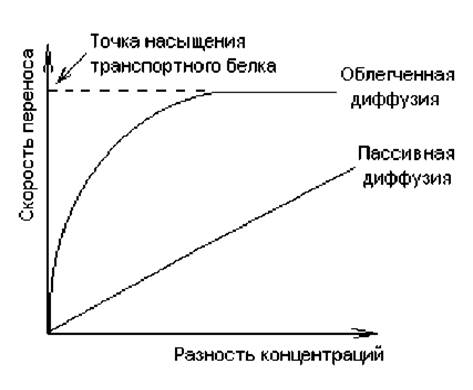
Рис. 4.10. Зависимость скорости процессов пассивной и облегченной диффузии от разности концентраций транспортируемого вещества
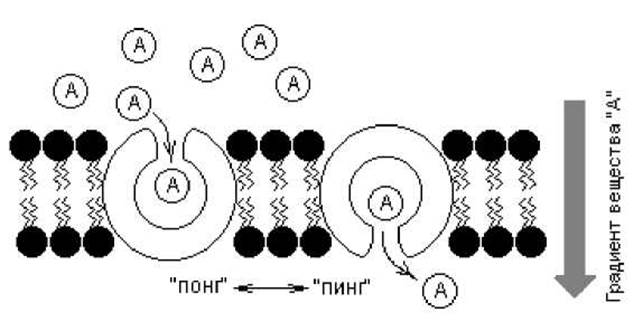
Рис. 4.11. Модель работы интегральных транспортных белков по типу «пинг-понг»
Работу каналов можно рассмотреть на примере ацетилхолинового рецептора. Этот интегральный белок находится в основном в мембранах нервномышечных соединений скелетных мышц. Он состоит из пяти субъединиц четырех типов и открывается в ответ на связывание ацетилхолина (нейромедиатор). При взаимодействии с ацетилхолином канал открывается, что связано с изменением конформации субъединиц, и пропускает определенные ионы (Na+, K+, Ca++ и некоторые другие), остается в таком положении 1 мс, а затем закрывается. Селективное перемещение катионов изменяет трансмембранный потенциал, в результате чего происходит электрическое возбуждение мышечной клетки, что приводит к сокращению мышцы. Изучение структуры ацетилхолинового рецептора показало, что пять белковых субъединиц встроены в бислой определенным образом: они организованы вокруг центральной поры диаметром 3 нм, через которую и транспортируются катионы. Непроницаемость канала для анионов и в три раза большую проницаемость для катионов, чем для незаряженных молекул, можно объяснить электростатическими взаимодействиями, возникающими благодаря присутствию в воротах канала биполярных или отрицательно заряженных групп.
Особым типом транспорта веществ в ходе облегченной диффузии является использование ионофоров, действие которых изучено на искусственных мембранах. Под ионофорами понимают низкомолекулярные вещества пептидной природы, избирательно транспортирующие через мембраны ионы. Различают ионофоры-каналообразователи (грамицидин А, амфотерицин В и др.) и ионофоры-переносчики (валиномицин, энниатины, боверицин).
Валиномицин представляет собой антибиотик депсипептидной природы, организованный по типу ионной «ловушки». В неполярных растворителях конформация валиномицина напоминает собой браслет, внутренняя полость которого точно подогнана под размеры ионов калия. Внешняя сфера валиномицина гидрофобна, в результате чего он способен перемещаться в липидном бислое и транспортировать через него ионы.
Хорошо изученным примером ионофоров-каналообразователей служит грамицидин А. Это антибиотик пептидной природы, состоящий из 15 аминокислот. Две молекулы грамицидина могут пронизывать мембрану в виде двойной спирали или образуя димер «голова к голове». В таких конформациях молекулы грамицидина А формируют полый цилиндр, по которому могут перемещаться ионы металлов.
В биологических мембранах ионофорный тип транспорта до сих пор не обнаружен.
Активный транспорт. Это сопряженный с потреблением энергии перенос молекул или ионов через мембрану против электрохимического градиента, в котором задействованы транспортные белки. Благодаря активному транспорту в жизнеспособных клетках между двумя сторонами мембраны поддерживается разность потенциалов, т. е. электрический заряд, при этом у большинства изученных клеток внутреннее содержимое заряжено отрицательно по отношению к внешней среде.
Активный транспорт сопряжен со значительными затратами энергии: некоторые клетки тратят более трети всей запасенной энергии для создания ионного градиента на мембране. Это необходимо для таких жизненно важных процессов, как осморегуляция, генерация и передача нервных импульсов, перенос в клетки питательных веществ (сахаров, аминокислот и др.).
Разнообразные системы активного транспорта отличаются друг от друга, в первую очередь тем, что служит для них источником энергии: АТР, ионный градиент, фосфоенолпируват, видимый свет. Наиболее хорошо изученной системой активного транспорта является натрий-калиевая (Na+/K+-АТР-аза, функционирующая в плазматических мембранах животных клеток. Этот интегральный белок состоит из двух субъединиц: бульшая представлена полипептидом, имеющим участки связывания для ионов натрия и АТР на цитоплазматической поверхности, а ионов калия — на наружной; меньшая субъединица является гликопротеином. Работа (Na+/K+)-АТР-азы заключается в следующем: при гидролизе одной молекулы АТР из клетки выкачивается 3 иона Na+, а извне в клетку проводится 2 иона K+, т. е. выводится больше положительных ионов, чем проводится внутрь клетки. Так на внутренней стороне мембраны создается избыточный отрицательный заряд, и клетка становится электрогенной. В мембранах обычно присутствуют проводящие пути для облегченной диффузии ионов натрия и калия по электрохимическому градиенту, и этот транспорт, хотя и с малой скоростью, совершается. Однако в жизнеспособной клетке не происходит уравнивания концентраций ионов, создающих электрохимический градиент на мембране, благодаря постоянной работе первичных активных переносчиков, таких, как (Na+/K+-АТР-аза. Таким образом, ионные насосы, принимающие участие в первичном активном транспорте, осуществляют перемещение заряда на мембране и создают на ней электрохимический градиент, в котором заключена энергия.
Вторичные активные переносчики используют электрохимические градиенты в качестве движущей силы для транспорта растворимых веществ. Этот процесс можно проследить на примере клеток эпителия кишечника. Образуемые в кишечнике при переваривании пищи строительные блоки (аминокислоты, глюкоза и др.) поступают в кровь при диффузии через мембраны кровеносных сосудов, и эта диффузия осуществляется в ходе симпорта (однонаправленного транспорта) с ионами натрия. Ионы натрия стремятся возвратиться в клетку согласно закономерностям облегченной диффузии и как бы тянут с собой молекулы питательных веществ. В мембранах обнаружены специфические переносчики сахаров и разных аминокислот, которые функционируют в системе активного транспорта, накапливая в клетке эти вещества, извлекая их даже из очень разбавленных растворов, т. е. против химического градиента. Эти же транспортные системы могут участвовать и в облегченной диффузии, если вещества транспортируются по химическому градиенту. Кроме описанного выше примера симпорта питательных веществ вместе с возвращающимися в клетку ионами натрия, существует и разнонаправленный транспорт — антипорт. Например, белок полосы 3 эритроцитов осуществляет сопряженный транспорт Cl- и HCO3- в противоположных направлениях через эритроцитарную мембрану.
У аэробных бактерий транспорт питательных веществ в клетку осуществляется в ходе симпорта не с ионами Na+, а с протонами. Наилучшим образом охарактеризованным примером подобного переносчика служит лактозопермеаза кишечной палочки. Этот интегральный белок использует протонный электрохимический градиент, созданный на мембране в результате окислительного фосфорилирования, для симпорта лактозы: с каждым возвращенным в клетку протоном переносится одна молекула лактозы.
Следует отметить, что АТР-азы представляют собой ферменты, катализирующие взаимообратимые реакции: при гидролизе АТР ионы транспортируются против электрохимического градиента, а перенос ионов по электрохимическому градиенту через каналы АТР-азы может запускать синтез АТР.
Эндоцитоз и экзоцитоз. Эти способы переноса веществ через мембраны связаны с образованием впячиваний (инвагинаций) мембраны и формированием особых мембранных везикул, обеспечивающих прохождение через мембрану крупных макромолекул и частиц. При этом эндоцитоз обеспечивает поглощение клеткой веществ, а экзоцитоз — выделение из клетки. Принято делить цитозы еще на два типа: пиноцитоз и фагоцитоз. Пиноцитоз — это механизм, с помощью которого через мембрану проводятся белки и другие макромолекулы в жидкой фазе. Фагоцитоз представляет собой поглощение клеткой крупных частиц, например бактерий, вирусов. Эти виды транспорта характерны в основном для эукариотических клеток, причем у животных фагоцитоз осуществляют только специализированные клетки, такие, например, как макрофаги. Для многих простейших, например амеб, фагоцитоз является основным способом питания.
Важной особенностью цитозов является последовательное образование и слияние везикул, в которых заключено транспортируемое вещество, причем секретируемые и поглащаемые молекулы локализуются в везикулах и не смешиваются с другими макромолекулами или органеллами клетки. С помощью не установленного пока механизма каждый пузырек сливается только со специфическими мембранными структурами (рис. 4.12).
В основе цитозов лежит еще одно характерное свойство липидных слоев биомембран — способность к агрегации, в результате чего мелкие везикулы объединяются в более крупные или происходит объединение везикул с плазматической мембраной клетки. Такой механизм основан на универсальности структуры биомембран, участвующих в формировании клеточных органелл и протопластов. Аналогичное явление можно наблюдать в пенах, где мыльные пузыри, также состоящие из амфифильных молекул (мыла — соли жирных кислот), обладают тенденцией к объединению с образованием более крупных структур. Способность мембран к агрегации лежит в основе такого широко распространенного способа переноса генетической информации, как слияние протопластов.
Скорость цитозов удивительно высокая. Показано, что клетки печени поглощают путем эндоцитоза за 1 ч количество жидкости, составляющее не менее 20% их объема, и количество мембранного материала, по площади превышающее в пять раз площадь их плазматической мембраны. Сходство цитозов с другими способами транспорта веществ через биомембраны состоит в том, что переносимое вещество должно быть «узнано» мембранными компонентами, иными словами, и в этом случае проявляется избирательная проницаемость мембран для различных соединений.
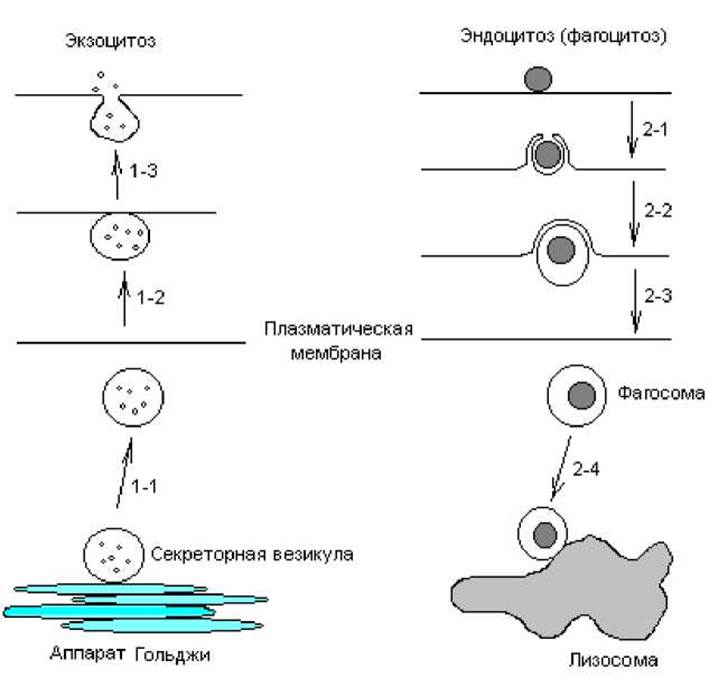
Рис. 4.12. Схематическое изображение цитозов. Стадии: 1-1 — от аппарата Гольджи отшнуровывается секреторная везикула; 1-2 — везикула диффундирует к плазматической мембране и слипается с ней; 1-3 — слияние везикулы с мембраной и выход содержимого везикулы в окружающую среду; 2-1 — частица обволакивается мембраной; 2-2 — фагосома отшнуровывается от мембраны; 2-3 — фагосома диффундирует к лизосоме; 2-4 — фагосома слипается с лизосомой и сливается с ней