Биохимия и молекулярная биология - Белясова Н.А. 2002
Метаболизм. Процессы, приводящие к запасанию энергии
Катаболические пути
Катаболизм углеводов
Основными источниками углерода и энергии, а значит, и пищевыми субстратами для большинства организмов (исключая растения) служат углеводы. Среди них на планете в наибольшей мере распространены целлюлоза, ее производные и крахмал. Кроме этих полисахаридов, большое значение имеют гликоген, инулин, хитин, ксиланы, пектиновые вещества и др. Большинство перечисленных полисахаридов расщепляется при участии специфических ферментов на моносахариды, среди которых преобладают гексозы и пентозы. Зачастую расщепление поли- и олигосахаридов осуществляется при участии ферментов фосфорилаз, и тогда образованные продукты оказываются фосфорилированными.
Особенностью сахаров является наличие при каждом атоме углерода атома кислорода, что делает возможным химическую атаку этих субстратов практически в любой точке молекулы. Кроме того, моносахариды и в первую очередь их фосфорилированные формы способны к изомеризации: карбонильные группы, атомы водорода могут легко перемещаться в соседнее положение или изменять свое пространственное положение в молекуле с помощью изомераз. Таким образом, появляется возможность перехода от любой гексозы или пентозы к любой другой, изомерной ей. По этой причине, несмотря на многообразие и сложность процессов обмена углеводов, можно выделить несколько типичных путей их превращения, в частности катаболизма, имеющих выраженные отличительные особенности. Такими путями служат: гликолиз, пентозофосфатные пути и путь Энтнера—Дудорова.
К закономерностям катаболизма моносахаридов относится обязательная начальная стадия активации свободных моноз, которая осуществляется в ходе фосфорилирования. В результате образуются фосфорные эфиры моносахаридов, способные вступать в дальнейшие превращения.
Гликолиз. Этот способ катаболизма сахаров называют иначе фруктозо-1,6-дифосфатным путем (по названию ключевого соединения) или путем Эмбдена—Мейергофа—Парнаса (по именам его исследователей).
Гликолиз считается наиболее универсальным и самым выгодным с энергетической точки зрения путем катаболизма гексоз. Процесс открыт в 1897 г. братьями Бухнерами, и его название происходит от двух греческих корней: glicos — сахар и lysis — растворять. В ходе гликолиза происходит не требующее участия молекулярного кислорода многоэтапное превращение гексоз в пируват, что сопровождается образованием АТР и восстановительных эквивалентов.
Реакции гликолитического пути осуществляются в цитозоле. Все промежуточные соединения имеют фосфорилированную форму. Для запасания энергии используется механизм субстратного фосфорилирования.
Превращение глюкозы в пируват (рис. 9.3) требует участия десяти ферментов и осуществляется в ходе следующих стадий: подготовка к разрыву гексозной цепи, разрыв цепи и образование глицеральдегид-3-фосфата, первое и второе субстратное фосфорилирование.
Процесс начинается с фосфорилирования глюкозы (активация молекулы) с участием АТР (донор фосфатной группы) и фермента гексокиназы. Образующийся глюкозо-6-фосфат в следующей реакции изомеризуется глюкозофосфатизомеразой во фруктозо-6-фосфат, который претерпевает второе фосфорилирование за счет АТР с образованием фруктозо-1,6-дифосфата.
Фруктозо-1,6-дифосфат является ключевым промежуточным продуктом гликолитического пути: именно данное соединение отличает гликолиз от других путей катаболизма сахаров. Кроме того, на уровне этого вещества осуществляется регуляция скорости всего процесса гликолиза. Активность катализирующего данную реакцию фермента (фосфофруктокиназы) ингибируется высокими концентрациями АТР, при этом снижается сродство фосфофруктокиназы к субстрату — фруктозо-6-фосфату. Кроме того, фосфофруктокиназа ингибируется цитратом — ранним промежуточным продуктом цикла трикарбоновых кислот. Известна и активация фосфофруктокиназы: ее осуществляют ADP и неорганический фосфат. Таким образом, фосфофруктокиназа наиболее активна в условиях, когда в клетке мало АТР (много ADP) и недостает строительных блоков.
На следующем этапе гликолиза фруктозодифосфат расщепляется с участием фруктозодифосфатальдолазы на два триозофосфата: дигидроксиацетонфосфат и глицеральдегидтрифосфат. Эти продукты представляют собой изомеры и легко переходят один в другой под действием триозофосфатизомеразы. Однако направление этой реакции сдвинуто в сторону образования глицеральдегид-3-фосфата, поскольку он все время выводится из реакционной смеси, претерпевая дальнейшие превращения. Глицеральдегид-3-фосфат служит субстратом первого субстратного фосфорилирования.
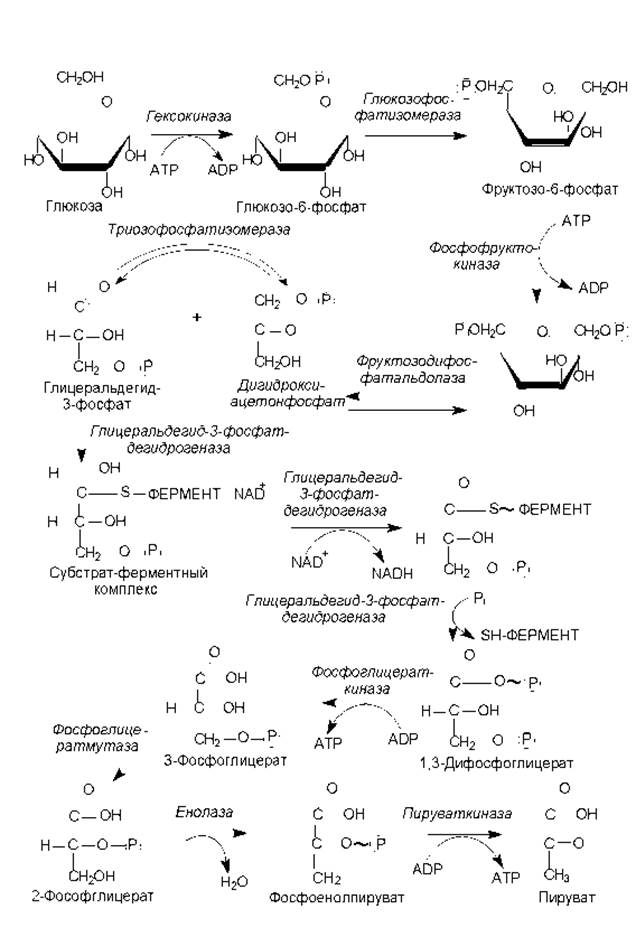
Рис. 9.3. Гликолитический путь катаболизма гексоз. Зигзагообразными линиями обозначены макроэргические химические связи
В ходе оставшихся этапов гликолиза, в отличие от предыдущих, энергия выделяется и запасается в форме АТР. Одной из подобных реакций является окисление глицеральдегид-3-фосфата. Фермент, катализирующий данную реакцию (глицеральдегид-3-фосфатдегидрогеназа), использует в качестве кофермента NAD и характеризуется содержанием большого количества сульфгидрильных групп (SH-групп). Окисление начинается со связывания глицеральдегид-3-фосфата с SH-группой фермента — формируется субстратферментный комплекс. Далее фермент катализирует перенос водорода с субстрата на NAD+, и восстановленный кофермент отделяется. Реакция дегидрирования и служит окислительной реакцией, поставляющей энергию: возникает комплекс фермента с ацильным остатком (тиоэфир), богатый энергией. Затем осуществляется фосфоролиз — перенос остатка глицеральдегид-3-фосфата совместно с макроэргической связью на фосфорную кислоту, что приводит к образованию 1,3-дифосфоглицериновой кислоты и исходной формы фермента. 1,3-Дифосфоглицерат представляет собой смешанный ангидрид фосфорной и карбоновой кислот и имеет высокий потенциал переноса фосфатной группы. Богатая энергией фосфатная группа переносится с 1,3-дифосфоглицерата на ADP с участием фосфоглицераткиназы и образованием продуктов: 3-фосфоглицерата и АТР. Так осуществляется первое субстратное фосфорилирование в гликолизе.
Заключительный этап гликолиза — второе субстратное фосфорилирование — начинается со внутримолекулярной перестройки, в ходе которой 3-фосфоглицерат изомеризуется в 2-фосфоглицерат (фермент фосфоглицератмутаза). 2-Фосфоглицерат дегидратируется в фосфоенолпируват при участии енолазы. В результате этой реакции образуется соединение, характеризующееся высоким потенциалом переноса фосфатной группы, таким образом, отщепление молекулы воды от 2-фосфоглицерата сопровождается перераспределением энергии внутри молекулы, и фосфатная связь у 2 атома углерода превращается из низко- в высокоэнергетическую. Последняя реакция гликолиза катализируется пируваткиназой, и в ходе нее фосфатная группа переносится на молекулу ADP и образуется конечный продукт пути — пируват.
Гликолитическое расщепление одной молекулы глюкозы приводит к образованию 4 молекул АТР (по две на каждую молекулу глицеральдегид-3-фосфата), из которых 2 расходуются на образование фруктозодифосфата. Таким образом, запасается всего 2 молекулы АТР. Кроме того, на одну молекулу глюкозы в этом процессе запасается 2 молекулы NADH (при окислении 2 молекул глицеральдегид-3-фосфата). Баланс гликолиза выглядит следующим образом:
1 С6Н12О6 → 2 С3Н4О3 + 2 АТР + 2NADH
Гликолитический процесс служит клетке для запасания энергии и восстановительных эквивалентов, а также является поставщиком «строительных блоков» в виде трехуглеродного, частично окисленного соединения — пировиноградной кислоты и некоторых промежуточных продуктов, в частности глицеральдегид-3-фосфата.
Пентозофосфатные пути. Эти пути катаболизма сахаров довольно разнообразны. В зависимости от условий и видовой принадлежности организма результатом данных процессов могут быть различные вещества. Осуществление реакций пентозофосфатных путей в обратном направлении используется клетками при фиксации СО2. Основной отличительной особенностью путей является образование в качестве промежуточных соединений пятиуглеродных сахаров, а также последовательное отщепление от 6-углеродного фосфорилированного сахара по одному атому углерода, который высвобождается в среду в виде СО2.
Пентозофосфатные пути (иначе, схема Варбурга—Диккенса—Хореккера, гексозомонофосфатный шунт, фосфоглюконатный путь) реализуются организмами реже, чем гликолиз, и обнаруживаются чаще всего в клетках бактерий.
После традиционной реакции активации глюкозы в ходе фосфорилирования образованный глюкозо-6-фосфат подвергается дегидрированию. Данную реакцию катализирует глюкозо-6-фосфатдегидрогеназа, а восстановительные эквиваленты акцептирует ее кофермент — NADP+. Образуется 6-фосфоглюконолактон (рис. 9.4). Данное соединение подвергается гидролизу (раскрытие кольца) с участием глюконолактоназы, а образованная 6-фосфоглюконовая кислота претерпевает второе дегидрирование (фосфоглюконатдегидрогеназа), после которого сразу же следует декарбоксилирование и образуется рибулозо-5-фосфат. Рибулозо-5-фосфат изомеризуется в два пятиуглеродных сахара — ксилулозо-5-фосфат и рибозо-5-фосфат (рис. 9.4).
Следующий этап пентозофосфатных путей представляет собой многократные межмолекулярные перестройки и изомеризацию промежуточных соединений, формирующихся из рибулозо-5-фосфата. Эти реакции направлены на образование 6-углеродного соединения (глюкозо-6-фосфата), которое может снова подвергнуться окислению и декарбоксилированию. В результате из 6 молекул рибулозо-5-фосфата получается 5 молекул глюкозо-6-фосфата (рис. 9.5).
Система структурной перестройки сахаров включает два фермента — транскетолазу и трансальдолазу, которые катализируют перенос двухуглеродных и трехуглеродных фрагментов, разрывая С—С-связи в двух положениях: по соседству с карбонильной (a) и у соседнего с карбонильной группой углеродного атома (ß) (на рис. 9.5 места, в которых связи подвергаются расщеплению, обозначены пунктирной линией).
При полном окислении глюкозы в пентозофосфатных путях молекула глицеральдегид-3-фосфата, образующаяся на конечном этапе, подвергается изомеризации с участием триозофосфатизомеразы в дигидроксиацетонфосфат, который затем вступает в реакцию альдольной конденсации со второй молекулой глицеральдегид-3-фосфата, и образуется фруктозодифосфат. Данная реакция является обратимой процессу расщепления фруктозодифосфата, которое имеет место в гликолизе (рис. 9.3), а фермент фруктозодифосфатальдолаза катализирует и прямое, и обратное превращения. Молекула фруктозо-1,6-дифосфата дефосфорилируется с образованием фруктозо-6-фосфата (фермент фосфатаза), а он, в свою очередь, изомеризуется в глюкозо-6-фосфат и возвращается к начальному этапу дегидрирования.
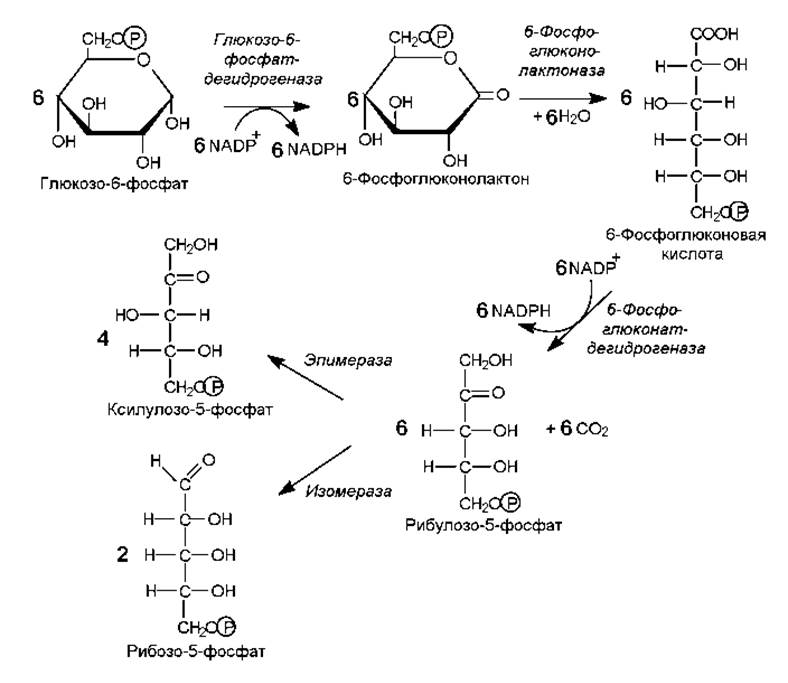
Рис. 9.4. Образование и изомеризация рибулозо-5-фосфата в ходе пентозофосфатных путей. Поскольку из одной молекулы гексозы может образоваться 6 молекул СО2, для удобства подсчета баланса процесс приведен на 6 молекул глюкозо-6-фосфата
Таким образом, на одну молекулу глюкозы при полном окислении в пентозофосфатных путях затрачивается 1 молекула АТР и образуется 6 молекул СО2 и 12 молекул NADPH. Таким образом, основной задачей пентозофосфатных путей является обеспечение клетки NADPH, который используется в процессах биосинтеза.
В ходе пентозофосфатных путей может и не происходить полного окисления глюкозы. В этом случае глицеральдегид-3-фосфат претерпевает иные превращения, например подвергается реакциям субстратного фосфорилирования под действием ферментов гликолитического пути.
Большое значение пентозофосфатные пути имеют для биосинтетических процессов: здесь образуются промежуточные соединения, играющие роль предшественников многих веществ. В частности, эритрозо-4-фосфат используется бактериями и растениями для синтеза ароматических аминокислот. Рибозо-5-фосфат служит субстратом для синтеза азотистых оснований и некоторых аминокислот.
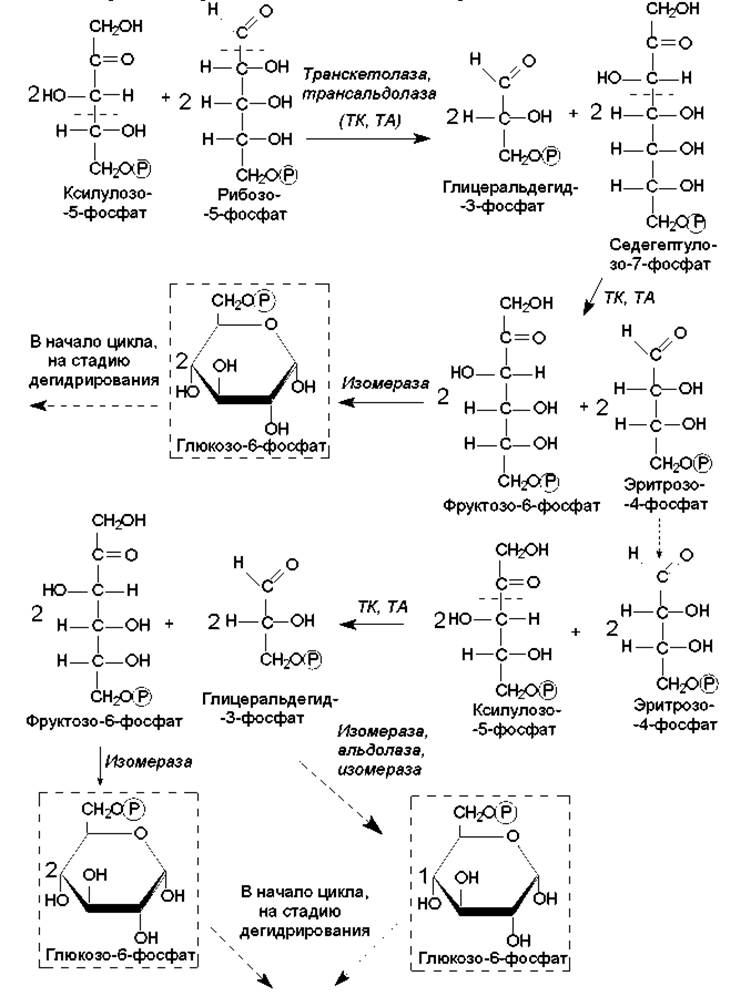
Рис. 9.5. Превращения изомеров рибулозо-5-фосфата в ходе пентозофосфатных путей при полном окислении глюкозы (в расчете на 6 молекул глюкозо-6-фосфата). Объяснения в тексте
Путь Энтнера—Дудорова (2-кето-3-дезокси-6-фосфоглюконатный). Этот процесс является еще одним способом катаболизма гексоз и представлен в основном в клетках микроорганизмов. Особое значение данный путь имеет для расщепления глюконата. Начальные стадии превращения глюкозо-6-фосфата совпадают с таковыми для пентозофосфатных путей, вплоть до образования 6-фосфоглюконовой кислоты. Далее, однако, 6-фофоглюконат подвергается а не окислению, а реакции дегидратации, с участием фермента 6-фосфоглюконат-дегидратазы. Образуется ключевое соединение данного пути — 2-кето-3-дезокси-6-фосфоглюко-нат. Стадия дегидратации осуществляется через образование промежуточного соединения (енола), который в результате таутомерного превращения переходит в 2-кето-3-дезокси-6-фосфоглюконат (рис. 9.6).
Далее альдолаза катализирует расщепление ключевого соединения на пируват и глицеральдегид-3-фосфат. Последний может вступать в гликолитический путь и подвергаться дальнейшим превращениям.
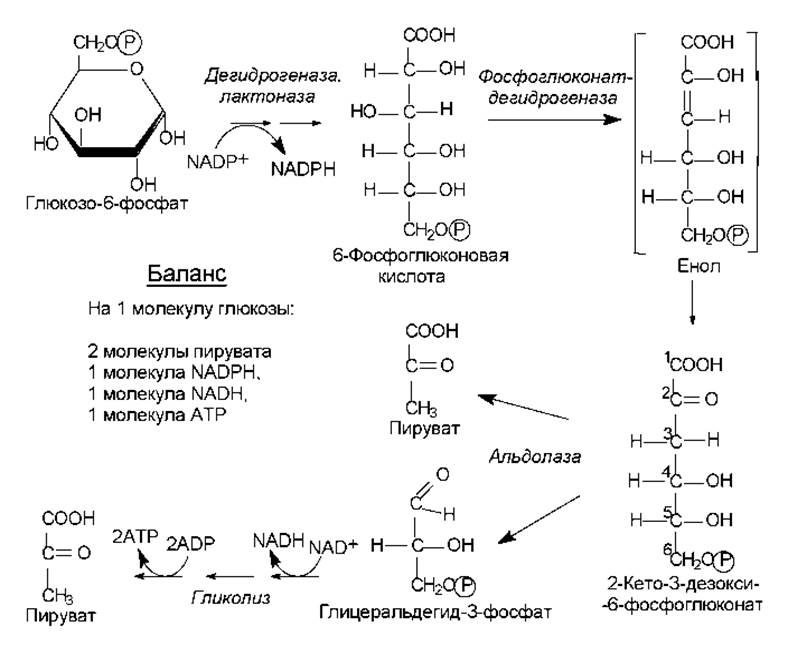
Рис. 9.6. Путь Энтнера—Дудорова и сопряженные с ним процессы
Как следует из баланса пути Энтнера—Дудорова, этот процесс менее выгоден с энергетической точки зрения, чем гликолиз: на молекулу глюкозы здесь запасается только 1 молекула АТР (из двух синтезированных одна тратится на фосфорилирование глюкозы).
Обзор основных катаболических путей позволяет увидеть, что расщепление «топливных» молекул при их неполном окислении сопровождается образованием двух основных метаболитов — пировиноградной кислоты и ацетил-СоА. Данные вещества могут использоваться как в биосинтетических путях в качестве «строительных блоков», так и подвергаться дальнейшим превращениям, обеспечивающим запасание клетками энергии. Поскольку в данном разделе обсуждаются процессы, приводящие к запасанию энергии, следует отметить, что судьба пирувата и ацетил-СоА зависит в первую очередь от наличия молекулярного кислорода в клетке, а также от ее ферментативного оснащения. Так, в клетках анаэробных микроорганизмов и тканей высших организмов пируват и ацетил-СоА могут подвергаться различного рода брожениям, завершающим процессы катаболического расщепления субстратов. В аэробных и факультативно-анаэробных клетках при наличии молекулярного кислорода пируват может подвергаться процессу окислительного декарбоксилирования, и образующийся ацетил-СоА вступает в цикл трикарбоновых кислот, где осуществляется его полное окисление до СО2 и Н2О, а формирующиеся восстановительные эквиваленты поступают в дыхательную цепь. Перечисленные процессы будут рассмотрены в следующих главах.