Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Стехиометрия и энергетика метаболических превращений
Стехиометрия клеточного роста и образования продуктов метаболизма
Материальный баланс по элементам и клеточный рост
Рост клеток и связанные с ним метаболические процессы можно описать такими же уравнениями, какие используются для изображения обычных химических реакций. Для этого прежде всего необходимо найти «химическую формулу» сухого клеточного вещества. Если известен элементарный состав данного штамма, растущего в определенных условиях (см., например, данные табл. 5.10), то не представляет труда найти и относительные значения коэффициентов в эмпирической формуле клетки CθHaOβNδ. Для того чтобы клетке отвечали одна строго определенная формула и соответствующая «молекулярная масса», удобно принять θ = 1 и затем в соответствии с элементным составом найти значения a, β и δ. Приведенные в табл. 5.10 «формулы» клеток получены именно таким путем. Один С-моль клеток определяют как количество клеток, в котором содержится один грамм-атом (12,011 г) углерода и которое отвечает эмпирической формуле клетки при θ = 1.
В качестве простейшего примера анализа стехиометрии клеточного роста на основе материального баланса по элементам рассмотрим прежде всего рост клеток в аэробных условиях, когда помимо новых клеток, СО2 и Н2О не образуется никаких иных продуктов метаболизма. Если источники углерода и азота мы обозначим формулами СНхОу и HlOmNn соответственно, то уравнение клеточного роста будет выглядеть следующим образом:
![]()
Здесь коэффициент при формуле клетки принят равным единице, что не отражается на общности уравнения, поскольку стехиометрические коэффициенты можно умножать или делить на любое одно и то же число.
Из уравнения (5.49) следует, что пять неизвестных стехиометрических коэффициентов a', b', с', d' и е' связаны четырьмя уравнениями:
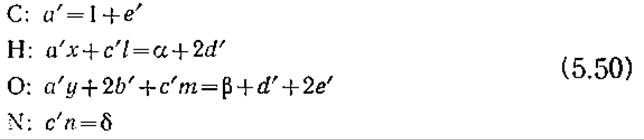
Еще одно уравнение можно получить, если экспериментальным путем определить дыхательный коэффициент (RQ) реакции клеточного роста. Дыхательным коэффициентом называют молярное (или объемное) отношение выделяемого при дыхании СО2
![]()
Для реакции роста клеток, выраженной уравнением (5.49),
RQ = e'/b' (5.52)
Таким образом, пять уравнений (5.52) и (5.50) содержат пять неизвестных стехиометрических коэффициентов, и если известен RQ, то можно определить все эти коэффициенты. Возможные затруднения, связанные с очень большими колебаниями значений стехиометрических коэффициентов при относительно небольших погрешностях в измерении RQ, мы рассмотрим в упражнении 5.16.
В общем случае химическая формула элементного состава клетки и стехиометрические коэффициенты в уравнении (5.49) зависят от состава питательной среды и условий, в которых находится клетка. Не следует забывать, что эта зависимость имеет описательный характер и в используемом здесь смысле отражает суммарный эффект множества независимых химических реакций. Чтобы получить какое-то представление о влиянии среды или условий клеточного роста на стехиометрию этого процесса и связать это влияние с обсужденными выше путями метаболизма, можно, например, разделить общую реакцию роста на несколько реакций, отвечающих различным метаболическим функциям (но не обязательно специфическим путям метаболизма).
Теперь мы несколько более детально рассмотрим стехиометрию роста хемогетеротрофного организма в аэробных условиях. С каждой реакцией мы будем связывать реакции генерирования и утилизации АТР. Ниже последние записаны отдельно и не включены в уравнения соответствующих реакций с участием соединении углерода, водорода, кислорода и азота из-за ряда практических затруднений, связанных со стехиометрией АТР; вкратце мы рассмотрим и эти затруднения.
Диссимиляция источника энергии:
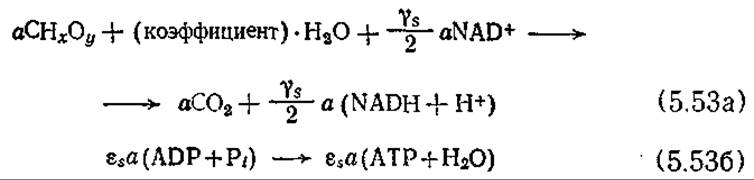
Окислительное фосфорилирование:
![]()
Биосинтез:

Поддержание и рассеяние энергии:
с(АТР + Н2О) → c(ADP + Pі) (5.56)
Взаимосвязь между этими реакциями можно установить, если допустить, что в рассматриваемых здесь условиях не накапливаются ни АТР, ни NADH, и, следовательно, синтез этих соединений должен уравновешиваться их утилизацией.
Приведенная выше усложненная, но более близкая к действительности система химических уравнений, отражающих клеточный рост, получена ценой резкого увеличения числа стехиометрических параметров. Заметьте, что в уравнениях (5.53) — (5.56), во-первых, стехиометрические коэффициенты отражают баланс по элементам в каждой отдельной реакции и, во-вторых, стехиометрический коэффициент биомассы CHаОβNδ принят равным единице. Поэтому коэффициенты, а, b и с в других реакциях отражают, по сути дела, степень протекания этих реакций относительно реакции роста клеток (биосинтеза). Стехиометрические коэффициенты для воды в двух случаях обозначены словом «коэффициент», а не каким-либо специальным символом, поскольку вовлечение воды в реакцию или ее выделение в результате реакции обычно практически не изменяет се общего количества в клетке. Символом εs обозначено число актов фосфорилирования в расчете на атом углерода, участвующий в процессах катаболизма.
Параметрами уВ, yS и yN обозначены степени восстановленности биомассы, источника углерода и источника азота соответственно. Степень восстановленности y соединения с брутто- формулой CHrOsNv определяется как у = 4 + r—2s—3v. Нетрудно видеть, что степень восстановленности СО2, Н2О и NH3 равна нулю.
Коэффициент YАTPmах обозначает массу клеток, образующихся в результате усвоения 1 моль АТР, при условии, что АТР расходуется только на цели биосинтеза. Предпринимались попытки оценить величину YATPmах по известным элементному составу клеток в путям биосинтеза [13]. Эти оценки показали, что, например, при росте Е. coli на глюкозе и неорганические солях YАTPmах составляет 28,8 г клеток на 1 грамм-моль АТР. Для перехода от единиц массы к мольным единицам в уравнение (5.556) включена молекулярная масса клеток, обозначенная символом MWB.
Как мы уже отмечали выше, P/О обозначает число актоз фосфорилирования ADP в расчете на один атом усвоенного кислорода; этот параметр характеризует эффективность окислительного фосфорилирования и в общем случае зависит от условий культивирования клеток. Расчетами на основе среднего элементного состава микроорганизмов показано, что теоретические максимальные значения P/О составляют 2,25, 2,50 и 3,00 при использовании в качестве основного питательного вещества ацетата, малата и глюкозы соответственно.
Прямое экспериментальное определение параметра P/О затрудняется тем обстоятельством, что изучать опытным путем отдельно пути синтеза или утилизации АТР крайне сложно; к тому же относительные количества образующегося и использующегося в каждом пути АТР зависят от условий культивирования клеток. Основная трудность здесь обусловлена частичной утилизацией АТР в процессах транспорта веществ через мембраны, в биосинтезе соединений, далее разлагаемых внутриклеточными гидролазами, и в других недостаточно изученных процессах (их иногда называют бесполезными циклами). Все перечисленные источники потребления АТР, сгруппированные здесь в одну «реакцию» (5.56) под общим названием «поддержание и рассеяние энергии», естественно, оказывают влияние на другие процессы генерирования и утилизации АТР.
Поскольку количество вновь синтезированного АТР должно быть равно количеству утилизированного АТР, то любые изменения в относительном количестве АТР, участвующего в «реакции» (5.56) (это количество отражает коэффициент с), повлекут за собой и изменения в количествах АТР, принимающего участие в других реакциях. Экспериментальные данные показывают, что потребность в АТР для поддержания клетки определяется не стехиометрическими факторами, а скоростями метаболических процессов, поэтому количественную оценку этих затрат мы отложим до изучения кинетики клеточного роста в гл. 7.
Здесь целесообразно упомянуть, что экспериментально найденные величины выхода биомассы в расчете па количество утилизированного АТР в анаэробных условиях обычно весьма мало зависят от типа субстрата и вида микроорганизма и в среднем величина YATP составляет 10,7 г клеточной массы на 1 моль АТР. У хемогетеротрофных организмов, как правило, значительно больше субстрата используется для выработки энергии, чем для роста биомассы. Так, в одной из работ было показано, что при росте пекарских дрожжей Saccharomyces cerevisiae в анаэробных условиях в богатой глюкозой среде (глюкоза являлась источником углерода) 98% усвоенного углерода расходовалось на выработку энергии и только 2% — на рост клеточной массы. Определить с достаточной точностью количество синтезируемого АТР в условиях аэробного роста оказалось крайне сложно. Имеющиеся данные говорят о том, что величина отношения P/О составляет от 0,5 до 1,8, т. е. значительно ниже теоретического значения. Изучение бактерии Aerobacter cloacae показало, что при ее росте в аэробной минимальной среде 55% глюкозы ассимилируется во вновь синтезированной биомассе, а 45% расходуется для выработки энергии. Эти данные свидетельствуют о том, что в аэробных условиях выход клеточной массы в расчете на количество утилизированного субстрата обычно выше, чем при анаэробном росте. Например, коэффициент Yх/s (масса клеток в граммах, образовавшихся из 1 моль глюкозы) Streptococcus faecalis в глюкозной среде составляет 21,5 в анаэробных условиях и 58,2 при аэробном росте В следующем разделе мы изучим влияние такого распределения субстрата на стехиометрию образования процессов метаболизма.