Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Молекулярная генетика и системы регуляции
Молекулярная генетика
Репликация ДНК и мутации
Поскольку ДНК содержит всю информацию, необходимую для развития и жизнедеятельности клетки, то последняя должна обладать практически безошибочно функционирующим механизмом копирования ДНК. При самовоспроизведении клеток каждая из дочерних клеток должна получить полную генетическую информацию в виде ДНК. Мы уже отмечали, что двуспиральная модель структуры ДНК обеспечивает точное воспроизных метаболитов. Отсюда следует, что перед технологом-биохимиком может возникнуть очень интересная и важная задача лементарная цепь, и в результате образуются две идентичные молекулы ДНК, каждая из которых содержит по одной цепи исходной ДНК. По-видимому, такая схема позволяет свести к минимуму ошибки в процессе репликации.
На самом деле процесс репликации ДНК осуществляется более сложным путем, чем это показано на рис. 6.11. Синтез ДНК представляет собой сложный ступенчатый процесс; особый фермент, называемый ДНК-полимеразой, легко строит дочерние цепи в направлении 5' → 3'. Этот процесс без затруднений происходит на одной из родительских цепей в направлении 3'→5'. На другой цепи ДНК-полимераза подбирает фрагменты дочерней цепи в направлении 5'→3' родительской цепи, а ковалентное связывание фрагментов осуществляет другой фермент — ДНК-лигаза. Таким образом происходит и синтез второй дочерней цепи в направлении 3'→5'.
Таблица 6.1. Число хромосом в клетках различных организмов в нормальном состоянии
Организм |
Число хромосом |
|
Прокариоты (гаплоидные) |
|
Бактерии |
1 |
|
Эукариоты (диплоидные) |
|
Клевер красный |
14 |
Пчела медоносная |
16 |
Пекарские дрожжи |
17 |
|
(Saccharomyces cerevisiae) |
|
Лягушка |
26 |
Гидра |
30 |
Кошка |
38 |
Крыса |
42 |
Человек |
46 |
Курица |
78 |
Здесь мы вкратце рассмотрим также различия в хранении информации и репликации между прокариотическими и эукариотическими ДНК. В нечетко отграниченном нуклеотиде прокариот имеется только одна хромосома (носитель генетической информации), содержащая кольцевую двойную спираль ДНК. Длина этой огромной молекулы составляет 1,2 мм, толщина около 20 ангстрем, а ее молекулярная масса равна примерно 2,8∙108. Этого достаточно для кодирования приблизительно 2000 различных белков. Кольцевая природа ДНК Е. coli впервые была выяснена в ходе генетических исследований, направленных на определение относительного расположения отдельных генов. Хромосома эукариот построена из молекулы ДНК, связанной с белками и, возможно, с небольшим количеством РНК.
Как показано в табл. 6.1, эукариотические клетки обычно содержат несколько хромосом. Некоторые эукариоты, например дрожжи, могут быть или гаплоидными (т. е. содержащими только по одной хромосоме каждого типа), или диплоидными, в каждой клетке которых содержится по две хромосомы каждого типа (за исключением, быть может, половых хромосом).
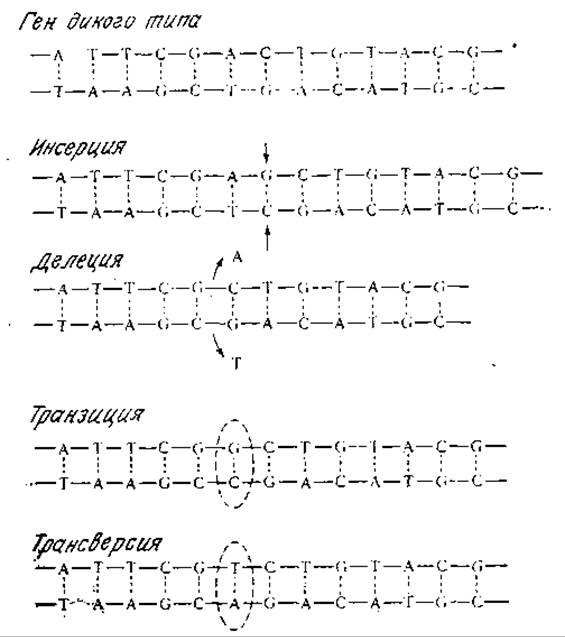
РИС. 6.12. Различные типы мутаций в последовательности пар оснований молекулы ДНК.
Мутацией называют изменение структуры ДНК, передающееся последующим поколениям. На молекулярном уровне мутация есть не что иное, как изменение нуклеотидной последовательности ДНК. Некоторые возможные варианты мутаций изображены на рис. 6.12. В какой-то степени мутация представляет собой спонтанный процесс, которому ДНК подвергается постоянно. Скорость спонтанных мутаций, однако, невелика и составляет в среднем 1 ошибку на миллион дупликаций гена.
Серьезность ошибки при копировании гена зависит от ее природы. При так называемой миссенс (изменяющей смысл) мутации кодон одной аминокислоты изменяется таким образом, что соответствующее место в аминокислотной последовательности белка занимает другая аминокислота. Этот тип мутации приводит, например, к аномальному гемоглобину, характерному для человека, страдающего серповидно-клеточной анемией; в этом конкретном случае в аномальном белке вместо Glu появляется остаток Val. Согласно генетическому коду (рис. 6.3), остаток Glu кодируют кодоны GAA и GAG, а валину соответствуют кодоны GUA и GUG. Отсюда следует, что замена всего лишь одного основания может приводить к тяжелому наследственному заболеванию. Другие изменения в кодонах могут приводить, например, к терминирующему кодону и тем самым вызывать преждевременное прекращение пептидного синтеза; мутации такого типа называются бессмысленными (нонсенс-мутациями).
Предлагалось несколько механизмов спонтанных мутаций. Во-первых, нуклеотидные основания ДНК могут существовать в нескольких структурно различных формах, называемых таутомерными. Предполагается, что изображенные на рис. 2.7 структуры доминируют в нативной ДНК; вместе с тем не исключается возможность и других таутомеров, образование пар оснований с которыми может приводить к ошибкам. Другая возможная причина спонтанных мутаций связана с функционированием ферментов, необходимых для синтеза и репарации (восстановления повреждений в структуре) ДНК. Наконец, некоторые промежуточные соединения в нормальном метаболизме клетки, например пероксиды, азотистая кислота, формальдегид, являются мутагенами, т. е. веществами, способными индуцировать химические мутации ДНК.
Действие химических мутагенов на ДНК широко изучалось путем выращивания клеток в среде, обогащенной такими агентами. К числу мутагенов относятся, например, соединения, структура которых близка структуре обычных оснований ДНК. В силу близости их строения и химических свойств в процессе биосинтеза ДНК в нуклеотидную цепь может быть включен не обычный нуклеотид, а его аналог. Другие типы мутагенов и способы проявления ими мутагенной активности приведены в табл. 6.2.
Второй общей причиной мутации является радиация. В частности, ДНК интенсивно поглощает ультрафиолетовое излучение, и при достаточной дозе это излучение довольно быстро убивает большинство клеток, а выжившие клетки подвергаются очень глубокой мутации. Любая клетка имеет в своем распоряжении ферментативные механизмы для репарации ДНК, поврежденной ультрафиолетовым излучением. Эти ферменты довольно сложным путем заменяют поврежденный сегмент, содержащий ковалентно-связанные пиримидиновые остатки.
В биохимической технологии явление мутации важно с нескольких точек зрения. Вспомните, например, рис. 1.10, на котором показано, как с помощью мутаций можно изменить природу микроорганизма, чтобы он стал более полезным для какой-либо определенной цели. Мутагены и ультрафиолетовое излучение часто используются для создания мутантов протестов, потенциально обладающих более высокой продуктивностью. В любой работе по изысканию новых штаммов микроорганизмов важно иметь в своем распоряжении эффективные способы идентификации и выделения мутантов, обладающих специфическими свойствами. В табл. 6.3 перечислены некоторые основные методы, применяющиеся для этой цели. Другие подходы к генетической трансформации продуцирующих организмов мы рассмотрим в разд. 6.2 и 6.3.
Таблица 6.2. Химические мутагены и соответствующие способы проявления мутагенной активностиa
Химический агент |
Мутагенный эффект |
Аналоги оснований |
Включение в ДНК вместо обычных оснований |
Азотистая кислота |
Дезаминирование пуриновых и пиримидиновых оснований ДНК |
Профлавин, акридиновый оранжевый |
Интеркалярное включение в стэкинг-комплексы оснований ДНК |
Алкилирующие агенты |
Модификация пуриновых оснований ДНК |
а Из монографии: Stanier R. Y., Doudoroff АТ, Adelberg E. A., The Microbial World, p. 418, Prentice-Hall, Inc., Englewood Cliffs, N. J., 1970.
С другой стороны, мутации могут создавать в биотехнологии и ряд затруднений. Для успеха микробиологического промышленного процесса часто необходимы чистые штаммы микроорганизмов, обладающие хорошо известными характеристиками. Вместе с тем никогда нельзя исключать возможность мутаций в таких культурах, поэтому необходима регулярная проверка их генетической гомогенности. Другие практические проблемы, связанные с мутациями и иными типами генетической нестабильности, мы рассмотрим в последующих главах.