Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Кинетика процессов утилизации субстрата, образования продуктов метаболизма и биомассы в культурах клеток
Кинетика образования продуктов метаболизма
Неструктурированные модели
Кинетика образования популяциями клеток продуктов метаболизма может быть описана аналогично росту клеточных популяций. Здесь также возможны структурированный и неструктурированный подходы. Как мы увидим в разд. 7.5.3, модели кинетики белкового синтеза можно разработать и на молекулярном уровне, если воспользоваться известными в настоящее время данными по регуляции молекулярных процессов.
Кинетика образования продуктов жизнедеятельности клеток наиболее проста в тех случаях, когда образование метаболитов и утилизация субстрата или клеточный рост связаны простым стехиометрическим отношением. Тогда скорость образования продуктов метаболизма можно выразить следующими уравнениями:
![]()
или
![]()
соответственно. Такие случаи типичны для микробиологических процессов типа I, рассмотренных в разд. 5.10.3. Примером может служить спиртовое брожение, кинетика которого в периодическом процессе отражена на рис. 7.29. Такого типа кинетику образования продуктов жизнедеятельности иногда называют сопряженной с ростом.
Во многих микробиологических процессах, особенно если речь идет о вторичных метаболитах, продукт жизнедеятельности клеток не образуется в сколько-нибудь значительных количествах в первых фазах периодического процесса вплоть до начала стационарной фазы или даже несколько позже, как, например, процесс биосинтеза пенициллина (рис. 7.30). Иногда в таких случаях кинетика образования продукта жизнедеятельности клеток удовлетворительно описывается простой, не сопряженной с ростом моделью, в которой скорость образования метаболита принимается пропорциональной не скорости роста клеток, а их концентрации.
В ставшей классической работе Льюдикина и Пайрета*, посвященной изучению молочнокислого брожения в присутствии бактерии Lactobacillus delbruekii, было показано, что в кинетику образования продукта метаболизма вносят вклад как сопряженные, так и не сопряженные с ростом факторы:
![]()
* Leudeking R., Piret Е. L., A Kinetic Study of the Lactic Acid Fermentation, J. Biochem. Microbiol. Technol. Eng., 1, 393 (1959).
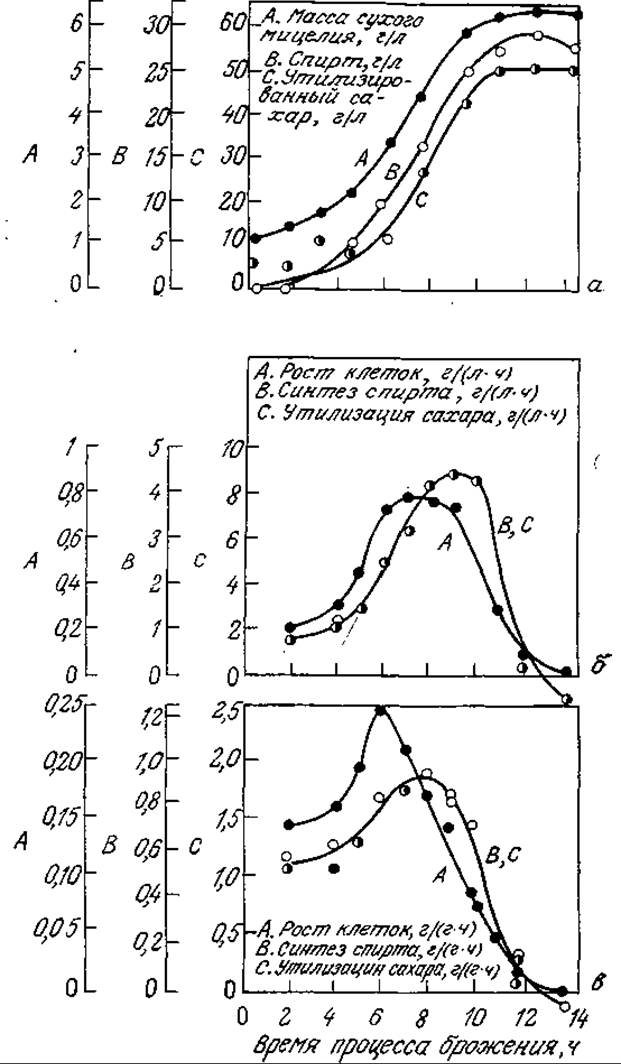
РИС. 7.29. Для процесса спиртового брожения характерна простая сопряженная с клеточным ростом кинетика образования продукта метаболизма. На рисунке указано изменение во времени биомассы, массы образовавшегося спирта и утилизированного сахара (а); объемных и удельных скоростей образования биомассы, утилизации субстрата и синтеза продукта метаболизма (спирта) (б, в). [Воспроизведено из работы: Leudeking R. Fermentation Process Kinetics, in Biochemical and Biological Engineering, Blakebrough N. (ed.), vol. 1, p. 203, Academic Press, Inc., (London) Ltd., London, 1967.]
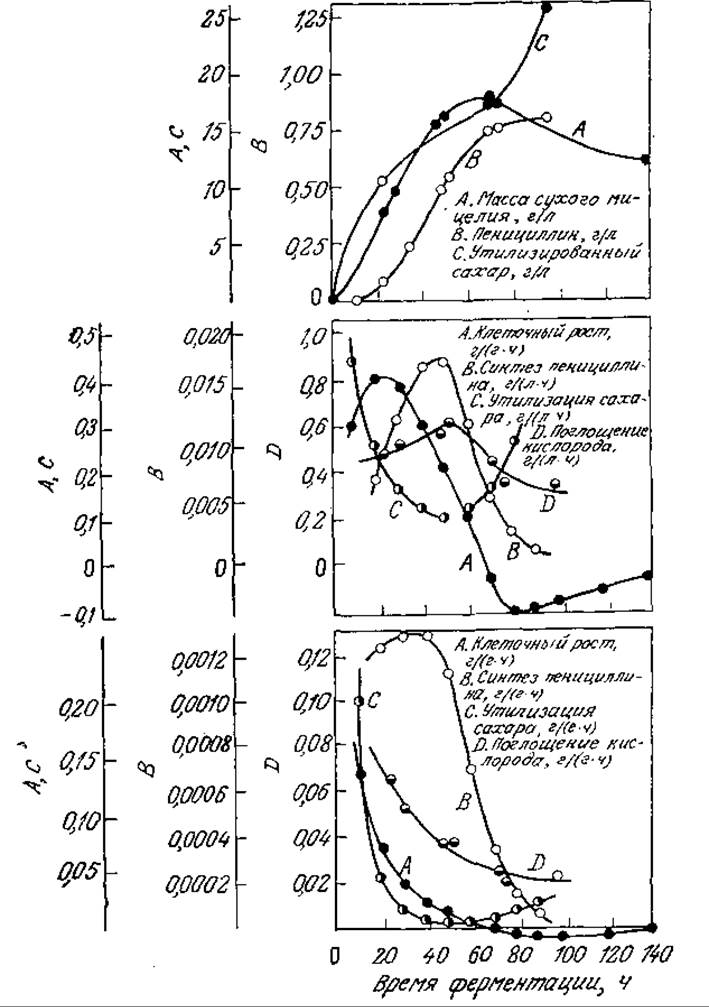
РИС. 7.30. Усложненная кинетика биосинтеза вторичного метаболита пенициллина. Указано изменение во времени биомассы, массы образовавшегося пенициллина и утилизированного сахара (а); объемных и удельных скоростей образования биомассы, синтеза пенициллина, утилизации сахара и кислорода (б, в). [Воспроизведено из работы: Leudeking R., Fermentation Process Kinetics, in Biochemical and Biological Engineering, Blakebrough N. (ed.), vol. I, p. 205, Academic Press, Inc., (London) Ltd., London, 1967.]
Такое уравнение скорости образования продукта метаболизма с двумя параметрами, часто называемое уравнением Льюдикина — Пайрета, оказалось чрезвычайно полезным при интерпретации экспериментальных результатов самых различных микробиологических процессов. Именно такую форму зависимости следует ожидать тогда, когда изучаемое вещество является конечным продуктом метаболического пути, связанного с выделением энергии (например, в некоторых процессах анаэробного брожения). В этих случаях первое и второе слагаемые правой части уравнения (7.93) можно рассматривать как меру энергии, расходуемой на клеточный рост и на поддержание клеток соответственно (см., например, работу [5]).
Пример 7.1. Последовательное определение параметров простого периодического процесса брожения. Рассмотрим модельный периодический процесс брожения, в котором клеточный рост описывается логистическим уравнением (7.51), а образование продукта метаболизма — уравнением Льюдикина — Пайрета (7.93):
![]()
Кинетику утилизации субстрата можно выразить следующим уравнением, в котором учтены превращение субстрата в клеточную массу и продукт метаболизма, а также его расход на поддержание клеток:
![]()
Подстановка уравнения (7П1.1) в (7П1.2) позволяет записать уравнение материального баланса по субстрату:
![]()
где
![]()
Эта и другие аналогичные модели позволяют описать многие практически важные микробиологические процессы, в том числе и такие, в которых образуется несколько конечных продуктов метаболизма [25]. К преимуществам этих моделей можно отнести и тот факт, что их параметры могут быть определены последовательно с помощью ряда специально разработанных удобных графических методов. Ниже мы вкратце рассмотрим эти методы.
Преобразованием уравнения (7.52) можно получить следующее выражение:
![]()
Нетрудно видеть, что определив экспериментально xs, можно затем по графику зависимости
![]()
Таблица 7П1.1. Значения параметров модели кинетики процессов биосинтеза внеклеточных полисахаридов различными микроорганизмамиa
|
Параметры образования продукта метаболизма |
||
Продукт процесса или микроорганизм |
ß |
а |
Ксантан Пуллулан (pH 4,5) Пуллулан (pH 5,5) Пуллулан (pH 6,5) Альгиновая кислота Pseudomonas sp. |
0,155 г продукта/г биомассы в час 0 0 0 0 10-3 г продукта/единиц биомассы в час |
1,83 г продукта/г биомассы 89% (по массе/(г в сутки в 100 мл) 135% (по массе)/(г в сутки в 100 мл) 110% (по массе)/(г в сутки в 100 мл) 1,60 г продукта/г биомассы 0 |
|
Продукт процесса или микроорганизм |
Параметры утилизации субстрата |
Параметры образования биомассы |
|
v |
η |
k, ч-1 |
|
Ксантан |
2,0 г субстрата/г биомассы |
0,284 г субстрата/г биомассы в час |
0,15 |
Пуллулан (pH 4,5) |
— |
— |
1,12 |
Пуллулан (pH 5,5) |
— |
— |
0,89 |
Пуллулан (pH 6,5) |
— |
— |
1,12 |
Альгиновая кислота |
6,6 г субстрата/г биомассы |
0,015 г субстрата/г биомассы в час |
0,12 |
Pseudomonas sp. |
0,165% (масса/объем)/единиц биомассы |
2,8∙10-2% (масса/объем)/единиц биомассы |
0,31 |
a Воспроизведено с разрешения из статьи: Ollis D. F., A Simple Batch Fermentation Model: Theme and Variations. Annals N. Y. Acad. Sci., 413, 144 (1983).
от t найти k (тангенс угла наклона прямой) и х0 (отрезок, отсекаемый прямой на оси ординат). Что касается параметров кинетики образования продукта жизнедеятельности клеток, то из уравнения Льюдикина — Пайрета для периодической культуры в стационарном состоянии следует
![]()
Интегрированием уравнения (7П1.1) при х, определяемом уравнением (7.51), получаем
![]()
Построив график зависимости левой части уравнения (7П1.7) от [x(t)—х0], можно определить а. Аналогичным путем можно найти параметры у и η в уравнении скорости утилизации субстрата.
В табл. 7П1.1 приведены найденные таким путем параметры кинетики четырех различных микробиологических процессов, в результате которых образуются внеклеточные полисахариды. Эти данные показывают, что процессы образования внеклеточных биополимеров по своим кинетическим характеристикам могут быть сопряжены с клеточным ростом (пуллулан и полиальгинат), не сопряжены с ним (биополимер из Pseudomonas sp.) или иметь смешанный характер (ксантан).
Таблица 7.4. Классификация микробиологических процессов по Дейндоерферу3
Тип процесса |
Описание |
Простой |
Превращение питательных веществ в продукты метаболизма осуществляется без накопления промежуточных веществ и характеризуется строго определенной стехиометрией |
Совместный |
Превращение питательных веществ в продукты метаболизма не сопровождается накоплением промежуточных веществ, но стехиометрия процесса может изменяться |
Последовательный |
Превращение питательных веществ в продукт метаболизма сопровождается накоплением промежуточного соединения |
Ступенчатый |
Превращению питательных веществ в продукт метаболизма предшествует полное превращение в промежуточное соединение, или питательные вещества селективно превращаются в продукт метаболизма в каком-либо определенном порядке |
а Deindoerfer F. И., Adv. Appl. Microbiol., 2, 321 (1960).
Временная зависимость концентрации продукта метаболизма в ходе периодического процесса может быть очень сложной, причем в среде могут накапливаться и подвергаться дальнeйшим превращениям несколько веществ. Различные возможные ситуации отражены в классификации микробиологических процессов, предложенной Дейндоерфером (табл. 7.4). В некоторых случаях сложная кинетика образования продукта жизнедеятельности клеток может отражать изменения в механизме метаболизма клетки. Показательным примером (важным как с точки зрения промышленного использования, так и в историческом аспекте) такой кинетики может служить синтез ацетона и бутанола бактерией Clostridium acetobutylicum (рис. 7.31). В первой фазе периодического процесса глюкоза превращается в уксусную и масляную кислоты, которые впоследствии перерабатываются (наряду с глюкозой) в ацетон и бутанол. Образование продуктов метаболизма клетки может сопровождаться также химическими превращениями метаболитов в среде, как, например, при спонтанном гидролизе пенициллина. Описание кинетики таких сложных процессов может потребовать включения этих дополнительных реакций в схему математической модели.
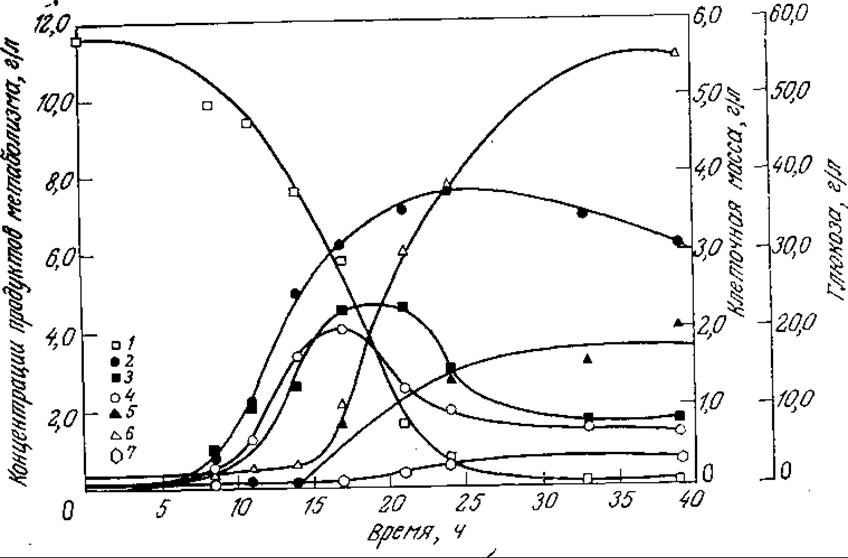
Рис. 7.31. Образование различных веществ в ходе периодического роста Cl. acetobulylicum при pH 5; приведены экспериментально найденные концентрации глюкозы (1), клеточной массы (2), уксусной кислоты (3), масляной кислоты (4), ацетона (5), бутанола (6), этанола (7). [Воспроизведено с разрешения из статьи: Costa J. М., Moreira A. R., Growth Inhibition Kinetics for the Acetone-Butanol Fermentation, ACS Symposium Series, 207, 501 (1983).]