ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 2. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ - 2014
ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
Метаболизм — это высококоординированная деятельность клетки, при которой происходит кооперативное взаимодействие многих мультиферментных систем (метаболические пути) для того, чтобы: (1) извлечь химическую энергию из окружающей среды (либо путем поглощения энергии солнечного света, либо при деградации богатых энергией питательных веществ); (2) превратить молекулы питательных веществ в собственные, характерные для клетки молекулы, включая предшественники макромолекул; (3) из предшественников-мономеров провести сборку макромолекул: белков, нуклеиновых кислот и полисахаридов; и (4) осуществить синтез и деграда
цию биомолекул, необходимых клетке для выполнения ее функций, — липидов мембран, внутриклеточных мессенджеров и пигментов.
Метаболизм включает сотни различных катализируемых ферментами реакций. В этой части книги (часть II) мы рассмотрим центральные метаболические пути, которые совсем не столь многочисленны и удивительно сходны у всех живых форм. Живые организмы можно разделить на две большие группы в зависимости от того, в виде каких молекул они получают углерод из окружающей среды. Автотрофы (такие как фотосинтезирующие бактерии и сосудистые растения) в качестве единственного источника углерода могут использовать диоксид углерода атмосферы, из которого они строят все свои углеродсодержащие биомолекулы (см. рис. 1-5). Некоторые автотрофные организмы, например, цианобактерии, могут также использовать атмосферный азот для того, чтобы производить все свои молекулы, которые содержат азот. Гетеротрофы не могут использовать углекислый газ атмосферы и должны получать углерод из окружающей среды в форме относительно сложных органических молекул, таких как глюкоза. Многоклеточные животные и большинство микроорганизмов — гетеротрофы. Автотрофные клетки и организмы относительно самодостаточны, в то время как гетеротрофные клетки и организмы для удовлетворения своих потребностей в углероде нуждаются в более сложных органических молекулах и поэтому должны питаться продуктами жизнедеятельности других организмов.
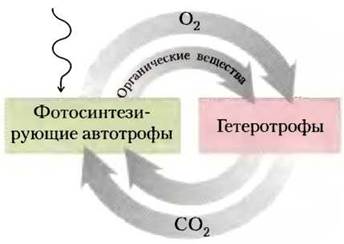
Многие автотрофные организмы используют фотосинтез, где в качестве источника энергии работает солнечный свет. Гетеротрофные организмы получают энергию при расщеплении органических питательных веществ, образуемых автотрофами. В биосфере автотрофы и гетеротрофы живут вместе в большом взаимозависимом цикле. Автотрофные организмы для построения своих органических биомолекул используют углекислый газ атмосферы. В этом процессе некоторые автотрофы генерируют кислород из воды. Гетеротрофы в качестве питательных веществ используют органические продукты, образуемые автотрофами, и выделяют в атмосферу углекислый газ. Для некоторых окислительных реакций с образованием диоксида углерода необходим кислород, который в реакциях окисления превращается в воду. Таким образом, между гетеротрофным и автотрофным естественными экосистемами постоянно происходит круговорот углерода, кислорода и воды. Движет этим глобальным процессом солнечная энергия (рис. 1).
Рис. 1. Круговорот углекислого газа и кислорода между автотрофным (фотосинтезирующим) и гетеротрофным доменами (экосистемами) биосферы. В этот круговорот вовлечены огромные массы веществ; биосферный оборот оценивается в ~4 • 1011 т углерода в год.
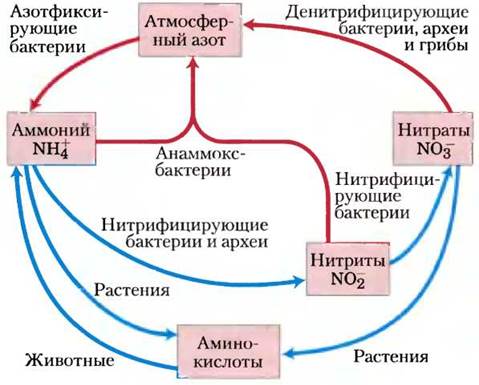
Все живые организмы нуждаются также в источнике азота, который необходим для синтеза аминокислот, нуклеотидов и других соединений. Растения в качестве источника азота используют главным образом аммиак или нитраты. Позвоночные животные должны получать азот в форме аминокислот или других органических соединений. Только некоторые организмы — цианобактерии и многие виды почвенных бактерий, обитающих в качестве симбионтов на корнях некоторых растений, — способны превращать (фиксировать) атмосферный азот N2 в аммиак. Другие бактерии (нитрифицирующие бактерии) окисляют аммиак до нитритов и нитратов, но есть и такие, которые превращают нитраты в свободный азот N2. Таким образом, в дополнение к глобальному круговороту углерода и кислорода в биосфере происходит круговорот азота, в котором участвует огромное количество этого элемента (рис. 2). Круговороты кислорода, углерода и азота, в которые в конечном счете вовлекаются все виды, зависит от естественного баланса между активностью продуцентов (автотрофов) и консументов (гетеротрофов) в нашей биосфере.
Рис. 2. Круговорот азота в биосфере. Газообразный азот N2 составляет 80% земной атмосферы.
Круговороты этих элементов приводятся в движение громадным потоком энергии, поступающим в биосферу извне, а затем преобразуемым в биосфере; все начинается с поглощения солнечной энергии фотосинтезирующими организмами с последующим использованием этой энергии для создания богатых энергией углеводов и других органических соединений. Эти питательные вещества выступают в качестве источника энергии для гетеротрофных организмов. В метаболических процессах и при любых превращениях энергии часть свободной энергии теряется путем выделения тепла в окружающую среду и увеличения энтропии системы. Таким образом, в биосфере материя участвует в непрекращающемся круговороте веществ, а энергия утилизируется — организмы не способны регенирировать энергию, которая рассеивается в виде теплоты и энтропии. Круговорот углерода, кислорода и азота осуществляется непрерывно, в то время как энергия постоянно превращается в форму, в которой она уже не может быть использована, — в тепловую энергию.
Метаболизм — совокупность всех химических превращений, которые происходят в клетке или организме и осуществляются посредством серии последовательных катализируемых ферментами реакций, называемых метаболическими путями. Реакции (стадии) метаболического пути следуют друг за другом в определенном порядке и на каждой стадии в систему привносится небольшое специфическое изменение ее химического состава. Обычно при этом происходит удаление, перемещение или добавление одного атома или функциональной группы. Превращение предшественника в конечный продукт идет через серию промежуточных продуктов метаболизма, называемых метаболитами. Термин промежуточный метаболизм часто применяют к совокупности ферментативных реакций всех метаболических путей, в которых происходит взаимное превращение предшественников, метаболитов и низкомолекулярных веществ (обычно с Мr <1000).
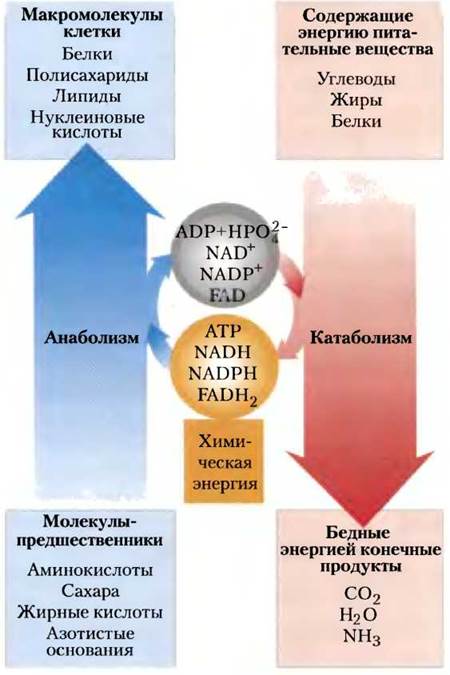
Катаболизм объединяет процессы деградации, при которых органические молекулы пищи (углеводы, жиры и белки) превращаются в низкомолекулярные и более простые конечные продукты (такие как молочная кислота, СО2, NH3). Катаболизм сопровождается высвобождением энергии, которая запасается в форме АТР и восстановленных переносчиков водорода (NADH, NADPH, FADH2). Остаток энергии рассеивается в виде тепла. Анаболизм, называемый также биосинтезом, включает процессы, при которых из мелких простых предшественников синтезируются более крупные и сложные молекулы, в том числе жиры, полисахариды, белки и нуклеиновые кислоты. Реакции анаболизма протекают с потреблением энергии, обычно получаемой при разрыве фосфатных связей АТР и восстановлении NADH, NADPH, FADH2 (рис. 3).
Рис. 3. Энергетическое сопряжение путей катаболизма и анаболизма. Катаболические пути поставляют химическую энергию в форме АТР, NADH, NADPH и FADH2. Эти переносчики энергии «работают» в анаболических путях, где происходят превращения низкомолекулярных веществ в макромолекулы.
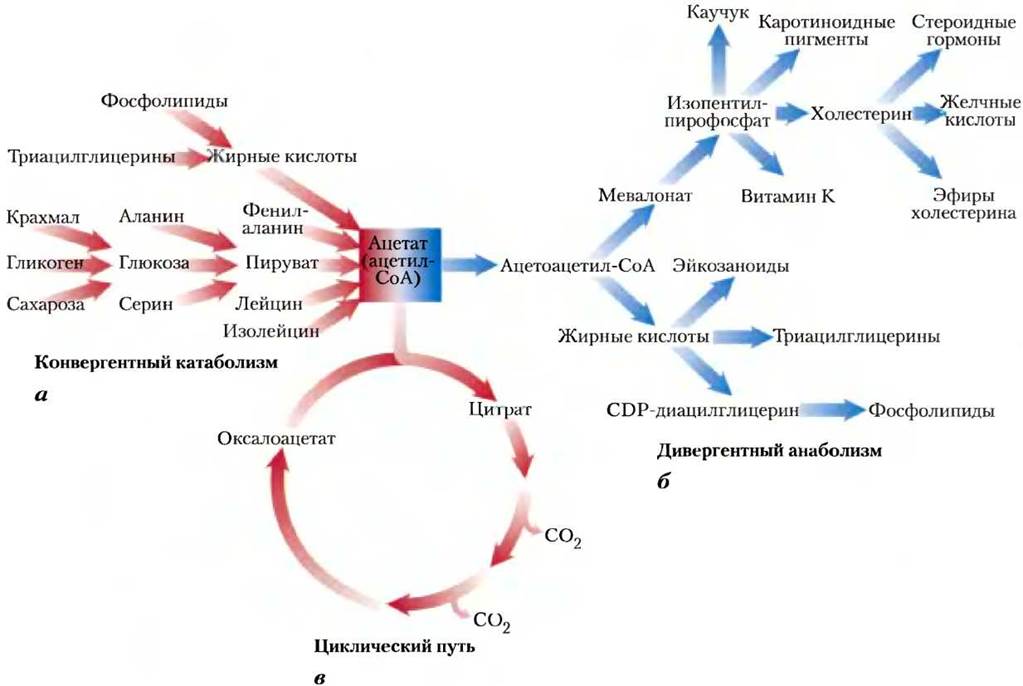
Одни метаболические пути представлены линейной цепочкой последовательных превращений. Другие метаболические пути имеют разветвления, т. е. из одного предшественника создается множество полезных конечных продуктов, или из нескольких исходных веществ образуется один продукт. Вообще, катаболические пути конвергентны, тогда как анаболические — дивергентны (рис. 4). Некоторые пути представляют собой реакционные циклы, когда молекула исходного вещества регенерируется в серии реакций, при этом несколько молекул, образованных из исходного реагента, вновь дают исходное вещество как продукт. В следующих
главах рассмотрены примеры различных метаболических путей.
Рис. 4. Три типа нелинейных метаболических путей, а — конвергентный катаболический путь; б — дивергентный анаболический путь; в — циклический путь, в котором одно из исходных веществ (в данном случае оксалоацетат) регенерируется, и цикл вновь начинается. Ацетат — ключевой промежуточный продукт метаболизма — возникает при распаде целого ряда богатых энергией молекул (а); он служит в качестве предшественника многих продуктов (б) и поглощается в катаболическом пути, известном как цикл лимонной кислоты (в).
У большинства клеток есть ферменты для проведения как реакций распада, так и реакций синтеза важных групп биомолекул, например, жирных кислот. Однако одновременное протекание синтеза и распада жирных кислот было бы расточительно, это предотвращается взаимной регуляцией анаболических и катаболических последовательностей реакций — когда одна реакционная цепочка активизируется, другая подавляется. Такая регуляция не могла бы осуществляться, если бы анаболические и катаболические пути катализировались одним и тем же набором ферментов, действующих в одном направлении для анаболизма и в противоположном — для катаболизма. Ингибирование фермента, вовлеченного в катаболизм, привело бы к ингибированию последовательности реакций анаболического направления. Катаболические и анаболические пути, которые имеют одинаковые концевые точки (например, глюкоза —>—> пируват и пируват —>—>глюкоза) могут использовать множество одинаковых ферментов. Но вот что обязательно — по крайней мере одна из стадий катаболического и анаболического путей катализируется разными ферментами и имеет различные механизмы регуляции; эти ферменты и являются местами отдельной регуляции. Более того, чтобы анаболический и катаболический пути были необратимыми, уникальные для каждого направления последовательности реакций должны включать хотя бы одну реакцию, которая термодинамически весьма благоприятна, другими словами, обратная ей реакция термодинамически невыгодна. Независимость регуляции катаболических и анаболических процессов усиливается и тем, что парные катаболический и анаболический пути обычно происходят в разных участках клетки. Например, катаболизм жирных кислот происходит в митохондриях, а синтез — в цитоплазме. Концентрации промежуточных метаболитов, ферментов и регуляторов могут поддерживаться в различных участках клетки на разных уровнях. Благодаря тому, что метаболические пути кинетически контролируются концентрацией субстрата, отдельные промежуточные продукты анаболизма и катаболизма тоже контролируют скорость метаболических процессов. Механизмам таких анаболических и катаболических процессов мы уделим особое внимание.
Метаболические пути регулируются на нескольких уровнях, как внутри клетки, так и внеклеточно. Наиболее быстро метаболические процессы реагируют на наличие субстрата. В общем случае внутриклеточная концентрация субстрата меньше Км; при этом скорость реакции определяется концентрацией субстрата (см. рис. 6-11). Второй способ внутриклеточно управлять скоростью метаболических процессов связан с аллостерической регуляцией (т. 1, с. 220) промежуточным продуктом метаболизма или коферментом, например, аминокислотой или АТР, которые сигнализируют о состоянии метаболизма внутри клетки. Когда клетка содержит достаточное для своих насущных потребностей количество, скажем, аспартата, или, когда уровень АТР в клетке такой, что дальнейшее потребление энергии в данный момент не нужно, эти сигналы аллостерически ингибируют активность одного или более ферментов в соответствующей последовательности реакций. У многоклеточных организмов метаболическая активность различных тканей регулируется и интегрируется ростовыми факторами и гормонами, которые действуют снаружи клетки. В некоторых случаях эта регуляция происходит фактически мгновенно (иногда быстрее, чем за миллисекунду) через изменения содержания внутриклеточных мессенджеров, которые изменяют активность ферментов путем аллостерической регуляции или их ковалентной модификации, например, при фосфорилировании. В других случаях внеклеточный сигнал приводит к изменению концентрации фермента в клетке, влияя на скорость его синтеза или распада. Такой эффект проявляется только через минуты или часы.
Часть II мы начинаем с описания основных энергетических закономерностей метаболизма (гл. 13). Затем обсудим главные пути катаболизма, по которым клетка получает энергию, окисляя разнообразные вещества (гл. 14-19). Подробно энергетические аспекты метаболизма рассмотрены в гл. 19. Она посвящена хемиосмотическому сопряжению энергии — универсальному механизму, по которому синтез АТР определяется трансмембранным электрохимическим потенциалом, возникающим либо в процессе окисления субстрата, либо в процессе поглощения солнечной энергии.
В гл. 20-22 рассматриваются главные анаболические пути, где за счет энергии АТР из более простых молекул-предшественников синтезируются углеводы, липиды, аминокислоты и нуклеотиды. В гл. 23 мы перейдем к детальному обсуждению метаболических путей у различных организмов, начиная с Echerichia coli и заканчивая человеком, и гормональных механизмов их регуляции и интеграции у млекопитающих.
И после этого мы, наконец, перейдем к изучению промежуточного метаболизма. Изучая клеточный метаболизм, надо помнить, что многочисленные реакции, описанные в этой книге, действительно происходят и играют решающую роль в живых организмах. Всегда старайтесь понять, какова роль того или иного химического процесса (реакции или всего метаболического пути) в организме. Каким образом изучаемые реакции связаны с другими реакциями, непрерывно происходящими в той же самой клетке с целью получения энергии и веществ, необходимых для поддержания жизнеспособности клетки? Как многоуровневые механизмы регуляции приводят в равновесие поглощение и выделение веществ и энергии, т. е. обеспечивают достижение стационарно-динамического состояния всего организма? Изучая метаболизм с таких позиций, вы испытаете захватывающее и вполне поучительное погружение в основы самой жизни, и полученные вами глубокие знания, безусловно, найдут множество применений в медицине, сельском хозяйстве и биотехнологиях.
Общая энергия вселенной остается постоянной; общая энтропия непрерывно увеличивается.
Рудольф Клаузиус, The Mechanical Theory of Heat with Its Applications to the Steam-Engine and to the Physical Properties of Bodies (Механическая теория тепла в приложении к паровому двигателю ифизическим свойствам тел), 1865
Благодаря изоморфизму энтропии и информации можно установить взаимосвязи между двумя формами энергии: энергией совершать действие и энергией направлять действие, которое совершается.
Франсуа Жакоб, La logique du vivant: une hitoire de I'heredite (Логика жизни: история наследственности), 1970