ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 2. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ - 2014
ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
14. ГЛИКОЛИЗ, ГЛЮКОНЕОГЕНЕЗ И ПЕНТОЗОФОСФАТНЫЙ ПУТЬ
14.5. Пентозофосфатный путь окисления глюкозы
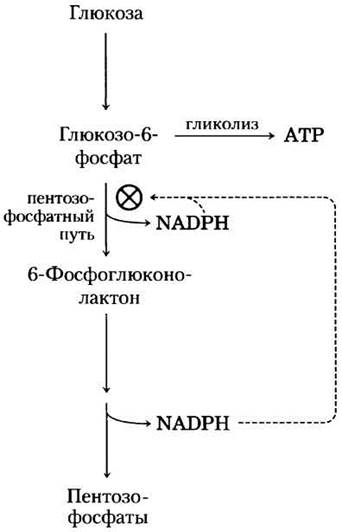
В большинстве тканей животных преобладающий путь катаболизма глюкозо-6- фосфата — это гликолитическое разложение до пирувата, основная часть которого затем окисляется в цикле лимонной кислоты, приводя в итоге к образованию АТР. Однако глюкозо-6-фосфат подвергается и другим превращениям до специальных продуктов, в которых нуждается клетка. В некоторых тканях наиболее важный процесс — окисление глюкозо-6-фосфата до пентозофосфата через пентозофосфатный путь (цикл или шунт), иначе называемый фосфоглюконатным или гексозомонофосфатным путем (рис. 14-20). На этом окислительном пути в качестве акцептора электронов выступает NADP+, превращающийся в NADPH. Быстроделящиеся клетки, такие как клетки костного мозга, кожи и слизистой кишечника, используют пентозы для синтеза РНК, ДНК и таких коферментов, как АТР, NАDН, FАDН2 и кофермент А.
Рис. 14-20. Общая схема пентозофосфатного пути. Образующийся на окислительном этапе NADРН используется для восстановления глутатиона (GSSG) (см. доп. 14-4) и поддержания восстановительного биосинтеза. На этом этапе также образуется рибозо-5-фосфат, необходимый для синтеза нуклеотидов, коферментов и нуклеиновых кислот. В тех клетках, которые не используют рибозо-5-фосфат для биосинтеза, на неокислительном этапе происходит превращение шести молекул пентозы в пять молекул гексозы (глюкозо-6-фосфата), что позволяет продолжать выработку NADРН и превращать глюкозо-6- фосфат в СO2 (за шесть циклов).
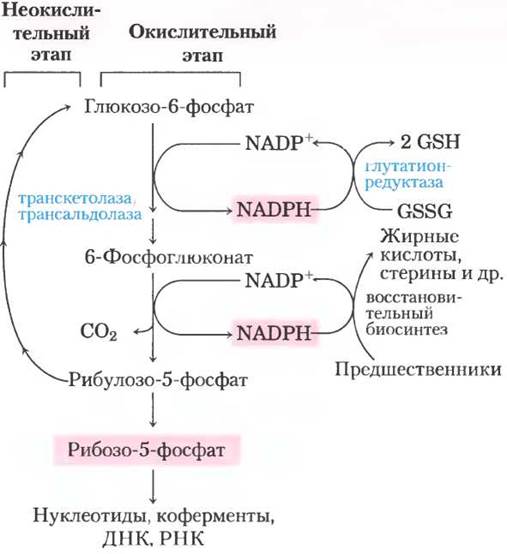
В других тканях важным продуктом пентозофосфатного пути являются не пентозы, а донор электронов NАDРН, необходимый для восстановительного биосинтеза и защиты от повреждающего действия радикалов кислорода. Наибольшую потребность в NАDРН испытывают те ткани, в которых происходит активный синтез жирных кислот (печень, жировая ткань, молочные железы) или холестерина и стероидных гормонов (печень, надпочечники, половые железы). Эритроциты, клетки хрусталика и роговицы подвержены непосредственному воздействию кислорода и, следовательно, вредному воздействию свободных радикалов, образуемых кислородом. Поддержание в этих клетках восстановительных условий (большие значения и глутатионOX/глутатионRED, и NADРН/NАDР+) позволяет предотвратить или ликвидировать окислительное повреждение белков, липидов и других чувствительных молекул. Образующийся в пентозофосфатном цикле NАDРН настолько важен для предотвращения окислительных повреждений в эритроцитах, что генетический дефект первого фермента данного метаболического пути глюкозо-6-фосфатдегидрогеназы может вызывать серьезные клинические последствия (доп. 14-4). ■
На окислительном этапе образуются пентозофосфаты и NADPH
Первой реакцией пентозофосфатного пути (рис. 14-21) является окисление глюкозо-6-фосфата до внутримолекулярного сложного эфира 6-фосфоглюконо-6-лактона под действием фермента глюкозо-6-фосфатдегидрогеназы (Г6ФДГ). Акцептором электронов выступает NADP+; равновесие реакции сильно смещено в сторону образования NADPH. Лактон гидролизуется под действием специфической лактоназы до 6-фосфоглюконата, который затем окисляется и декарбоксилируется под действием 6-фосфоглюконатдегидрогеназы с образованием кетопентозы рибулозо-5-фосфата. В этой реакции образуется вторая молекула NADPH. (Рибулозо-5-фосфат необходим для регуляции гликолиза и глюконеогенеза, что будет показано в гл. 15.) Далее фосфопентозоизомераза превращает рибулозо-5-фосфат в его изомер — рибозо- 5-фосфат. В некоторых тканях пентозофосфатный путь на этом завершается; общее уравнение процесса выглядит следующим образом:
Глюкозо-6-фосфат + NADP+ + Н2O —> рибозо-5-фосфат + СO2 + 2 NADPH + 2 Н+
Рис. 14-21. Окислительные реакции пентозофосфатного пути. Конечными продуктами данного метаболического пути являются рибозо-5-фосфат, СO2 и NADРН.
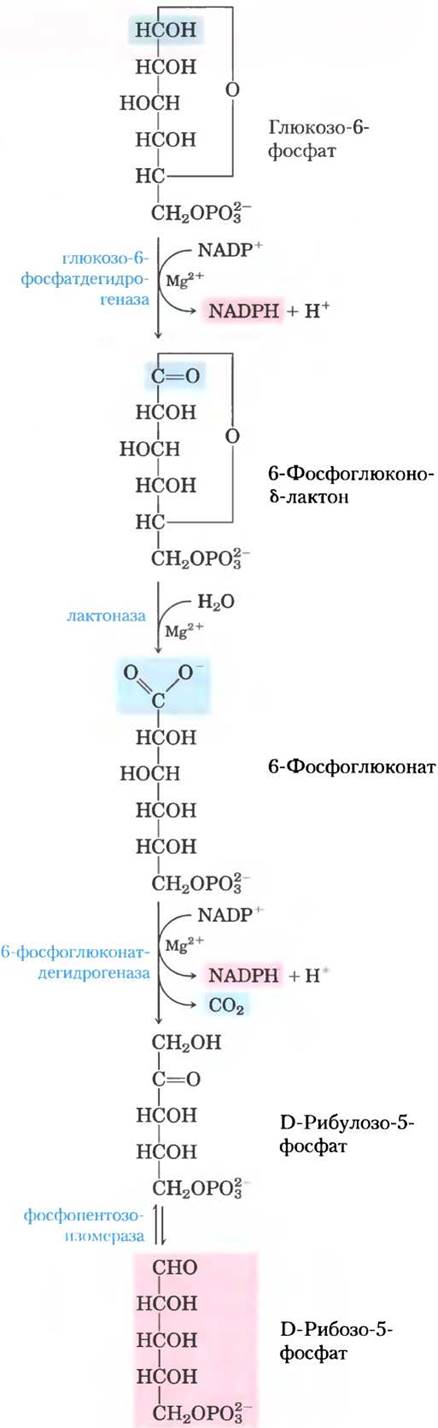
В результате образуется NADPH — восстановитель в реакциях биосинтеза, а также рибозо-5-фосфат — предшественник для синтеза нуклеотидов.
На неокислительном этапе пентозофосфаты превращаются в глюкозо-6-фосфат
В тканях, где особенно требуется NADPH, образующиеся на окислительном этапе пентозофосфатного пути пентозофосфаты вновь превращаются в глюкозо-6-фосфат. В этом неокислительном процессе рибулозо-5-фосфат сначала превращается в свой эпимер ксилулозо-5-фосфат:
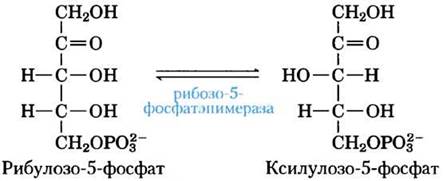
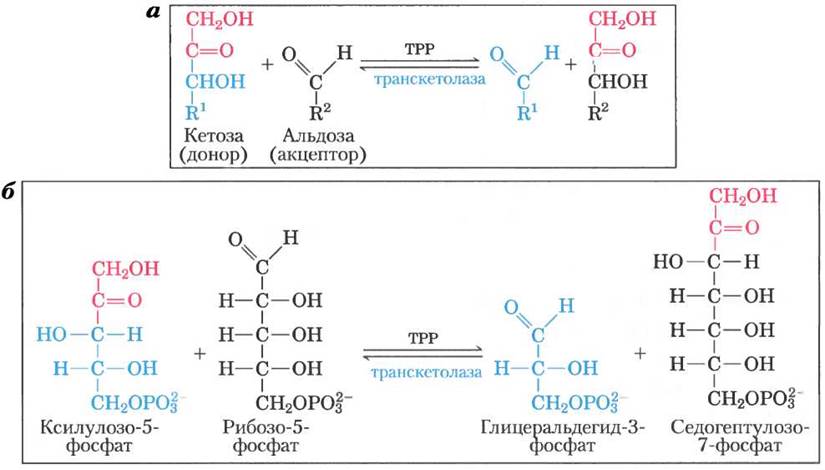
Затем в результате перегруппировки углеродного скелета (рис. 14-22) шесть пятиуглеродных фосфатов превращаются в пять шестиуглеродных фосфатов, завершая тем самым цикл и способствуя окислению глюкозо-6-фосфата с образованием NADPH. Реализация данного цикла приводит в итоге к превращению глюкозо-6-фосфата в шесть молекул СО2. В этом процессе участвуют два фермента, задействованные исключительно в пентозофосфатном пути, а именно, транскетолаза и трансальдолаза. Транскетолаза катализирует перенос двухуглеродного фрагмента от кетозы (донор) на альдозу (акцептор) (рис. 14-23, а). Первой реакцией пентозофосфатного пути с участием транскетолазы является перенос атомов С-1 и С-2 от ксилулозо-5-фосфата на рибозо-5-фосфат, в результате чего образуется семиуглеродный продукт седогептулозо-7- фосфат (рис. 14-23, б). Оставшийся от ксилулозы трехуглеродный фрагмент превращается в глицеральдегид-3-фосфат.
Рис. 14-22. Неокислительные реакции пентозофосфатного пути, а — в этих реакциях происходит превращение пентозофосфатов в гексозофосфаты, что позволяет непрерывно осуществлять реакции окислительного этапа пентозофосфатного пути (рис. 14-21). Здесь действуют как специфические ферменты (транскетолаза и трансальдолаза), так и ферменты, задействованные в реакциях гликолиза и глюконеогенеза. б — схема превращений шести пентоз (5С) в пять гексоз (6С). Обратите внимание, что реакции состоят из серии двух взаимопревращений (а). Каждая приведенная здесь реакция обратима; однонаправленные стрелки указывают направление реакций при непрерывном окислении глюкозо-6-фосфата. В темновых реакциях фотосинтеза эти реакции протекают в противоположном направлении (см. рис. 20-10).
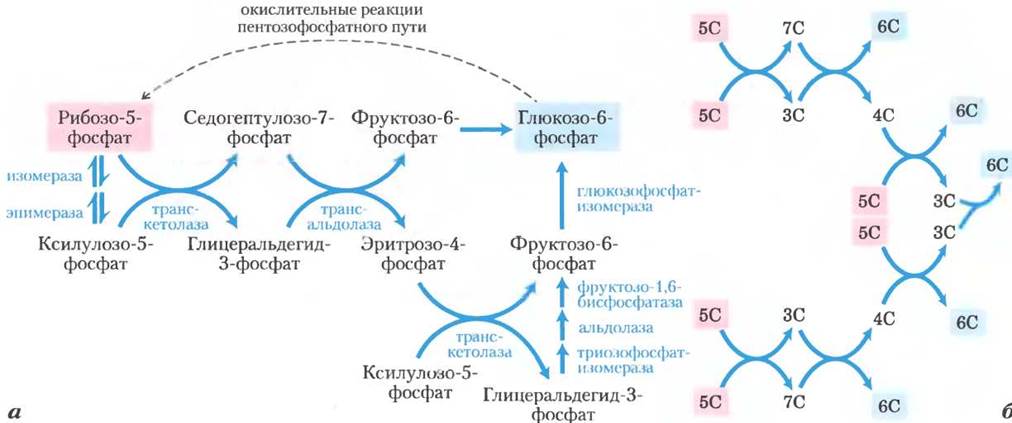
Рис. 14-23. Первая реакция пентозофосфатного пути, катализируемая транскетолазой. а — реакция, катализируемая транскетолазой, состоит в переносе двухуглеродного остатка от кетозы (донор) на альдозу (акцептор) с помощью ТРР-связанного фермента. б — превращение двух пентозофосфатов в триозофосфат и семиуглеродное соединение седогептулозо- 7-фосфат.
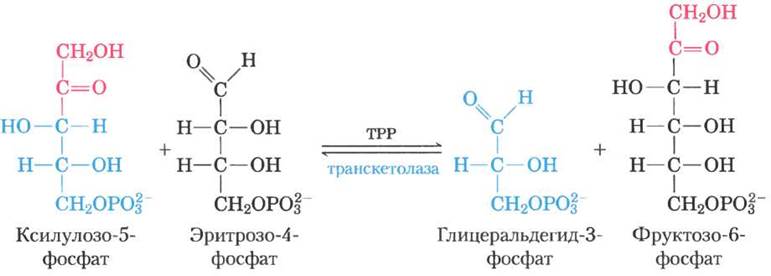
Далее фермент трансальдолаза катализирует реакцию, напоминающую альдолаз- ную реакцию гликолиза: происходит перенос трехуглеродного фрагмента от седогептулозо- 7-фосфата на глицеральдегид-3-фосфат, в результате чего образуются фруктозо-6-фосфат и тетроза эритрозо-4-фосфат (рис. 14-24). Теперь опять начинает действовать транскетолаза, которая из эритрозо-4-фосфата и ксилулозо- 5-фосфата образует фруктозо-6-фосфат и глицеральдегид-3-фосфат (рис. 14-25). Две молекулы глицеральдегид-3-фосфата, образовавшиеся при повторении этих реакций, превращаются в молекулу фруктозо-1,6-бисфосфата, как в процессе глюконеогенеза (рис. 14-16). И, наконец, фруктозобисфосфатаза-1 и глю- козофосфатизомераза превращают фруктозо-1, 6-бисфосфат в глюкозо-6-фосфат. Цикл завершен: шесть пентозофосфатов превращены в пять гексозофосфатов (рис. 14-22, б).
Рис. 14-24. Реакция, катализируемая трансальдолазой.
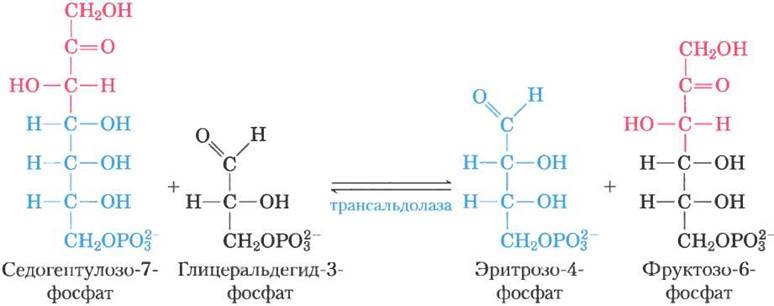
Рис. 14-25. Вторая реакция, катализируемая транскетолазой
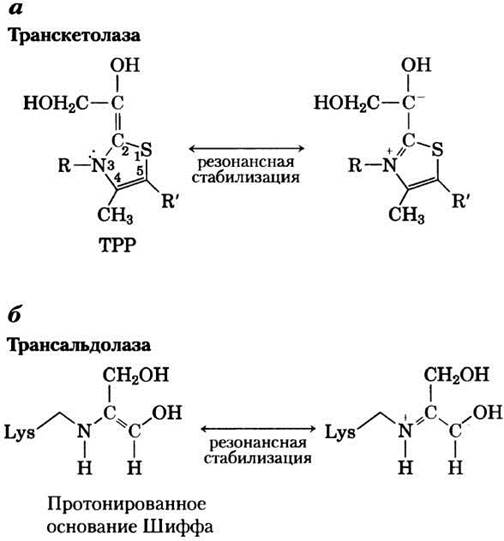
Для действия транскетолазы необходим кофактор тиаминпирофосфат (ТРР), который стабилизирует образующийся в реакции двухуглеродный карбанион (рис. 14-26, а), точно так же, как это было описано в случае пируватдекарбоксилазы (рис. 14-14). Трансальдолаза (рис. 14-26,6) стабилизирует карбанион, возникающий при образовании основания Шиффа между боковой цепью остатка лизина и карбонильной группой субстрата (кетозы).
Рис. 14-26. Стабилизация карбанионов путем ковалентных взаимодействий с транскетолазой и трансальдолазой. а — кольцо ТРР стабилизирует двухуглеродный карбанион, переносимый транскетолазой (механизм действия ТРР см. на рис. 14-14). б — в реакции, катализируемой трансальдолазой, стабилизация трехуглеродного карбаниона, возникшего в результате альдольного расщепления, происходит путем образования протонированного основания Шиффа между ε-аминогруппой боковой цепи лизина и субстратом.
Синдром Вернике-Корсакова усугубляется дефектом транскетолазы
Синдром Вернике-Корсакова связан с недостаточностью тиамина, который входит в состав тиаминпирофосфата (ТРР). Этот синдром чаще встречается при хроническом злоупотреблении алкоголем, который затрудняет всасывание тиамина в кишечнике. Симптомы заболевания могут усиливаться при мутации гена транскетолазы, что приводит к образованию фермента, сродство которого к ТРР может быть в 10 раз ниже, чем у нормального фермента. Этот дефект делает человека гораздо более чувствительным к недостаточности тиамина: даже умеренная недостаточность этого вещества (неощутимая для людей с нормальной формой транскетолазы) может привести к уменьшению содержания ТРР ниже того уровня, который необходим для насыщения фермента. В результате замедляются реакции пентозофосфатного пути в целом. У страдающих синдромом Вернике-Корсакова это приводит к усилению симптомов заболеваания, что выражается в потере памяти, спутанности сознания и частичном параличе. ■
Изображенный на рис. 14-21 процесс называют окислительным пентозофосфатным путем. Первые две реакции окисления характеризуются большим отрицательным изменением стандартной свободной энергии и необратимы во внутриклеточных условиях. Реакции неокислительного этапа пентозофосфатного пути (рис. 14-22) легко обратимы и, следовательно, предоставляют возможность для превращения гексозофосфатов в пентозофосфаты. Как мы увидим в гл. 20, превращение гексозофосфатов в пентозофосфаты занимает центральное место в фотосинтетической ассимиляции СО2 у растений. Данный путь метаболизма, называемый восстановительным пентозофосфатным путем, фактически представляет собой обращение процесса, изображенного на рис. 14-22, и использует многие из указанных ферментов.
Все ферменты, принимающие участие в пентозофосфатном пути, локализованы в цитозоле, подобно ферментам гликолиза и большинству ферментов глюконеогенеза. Действительно, все три процесса связаны наличием нескольких общих интермедиатов и ферментов. Глицеральдегид-3-фосфат, образующийся при действии транскетолазы, легко превращается в дигидроксиацетонфосфат под действием гликолитического фермента триозофосфатизомеразы, а две названные триозы под действием альдолазы образуют фруктозо-1,6-бисфосфат, как в процессе глюконеогенеза. Триозофосфа- ты же могут окисляться до пирувата в реакциях гликолиза. Путь превращения триоз определяется потребностью клетки в пентозофосфатах, NАDРН и АТР.
Глюкозо-6-фосфат распределяется между гликолизом и пентозофосфатным путем
Участие глюкозо-6-фосфата в гликолизе или пентозофосфатном пути зависит от текущих нужд клетки и концентрации NАDР+ в цитозоле. Без этого акцептора электронов первая реакция пентозофосфатного пути (катализируемая Г6ФДГ) протекать не может. Если в клетке в результате восстановительных реакций биосинтеза NАDРН быстро превращается в NАDР+, уровень NАDР+ повышается, происходит аллостерическая регуляция Г6ФДГ, и в итоге увеличивается количество глюкозо-6- фосфата, включающегося в пентозофосфатный путь (рис. 14-27). При снижении потребностей в NАDРН уменьшается уровень NАDР+, пентозофосфатный путь замедляется и глюкозоб-фосфат в большей степени направляется на гликолиз.
Рис. 14-27. Роль NАDРН в регуляции распределения глюкозо-6-фосфата между гликолизом и пентозофосфатным путем. Если NADРН образуется быстрее, чем используется в реакциях биосинтеза и для восстановления глутатиона (см. рис. 14-20), концентрация NADРН увеличивается и происходит ингибирование первого фермента пентозофосфатного пути. В результате больше глюкозоб-фосфата оказывается доступным для гликолиза.
Краткое содержание раздела 14.5 Пентозофосфатный путь окисления глюкозы
■ Окислительный пентозофосфатный путь (иначе называемый фосфоглюконатным или гексозомонофосфатным путем) состоит в окислении и декарбоксилировании глюкозо-6-фосфата у атома С-1 с восстановлением NАDР+ до NАDРН и образованием пентозофосфатов.
■ NАDРН служит восстановителем в реакциях биосинтеза, а рибозо-5-фосфат — предшественник нуклеотидов и нуклеиновых кислот. Быстрорастущие ткани, а также те клетки, что осуществляют активный биосинтез жирных кислот, холестерина или стероидных гормонов, направляют на пентозофосфатный путь больше глюкозо-6-фосфата, чем ткани, менее нуждающиеся в пентозофосфатах или восстановителе.
■ Окислительный этап пентозофосфатного пути представляет собой двухступенчатое превращение глюкозо-6-фосфата до рибулозо-5- фосфата, сопровождающееся восстановлением NАDР+ до NАDРН. Неокислительный этап состоит в превращении пентозофосфатов в глюкозо-6-фосфат, что запускает цикл заново.
■ На втором (неокислительном) этапе пентозофосфатного пути транскетолаза (с ТРР в качестве кофактора) и трансальдолаза катализируют взаимопревращения трех-, четырех-, пяти-, шести- и семиуглеродных сахаров, что приводит к обратимому образованию пяти гексозофосфатов из шести пентозофосфатов. В темновых реакциях фотосинтеза эти же ферменты катализируют обратный процесс, называемый восстановительным пентозофосфатным путем, а именно, превращение пяти гексозофосфатов в шесть пентозофосфатов.
■ Генетический дефект транскетолазы, снижающий сродство фермента к ТРР, усугубляет синдром Вернике-Корсакова.
■ Участие глюкозо-6-фосфата в гликолизе или пентозофосфатном пути в большой степени определяется отношением концентраций NАDР+ и NАDРН.