Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Биосинтез углеводов в животных тканях
Обходный путь требуется для превращения пирувата в фосфоенолпируват
Первая обходная реакция в глюконеогенезе - это превращение пирувата в фосфоенолпируват (рис. 20-2). Она не может быть простым обращением пируват- киназной реакции (разд. 15.7, д).

которая характеризуется большой отрицательной величиной изменения стандартной свободной энергии и потому в интактной клетке необратима. Вместо этого фосфорилирование пирувата достигается обходным путем - в последовательности реакций, которые у некоторых животных требуют совместного действия цитозольных и митохондриальных ферментов клеток печени (рис. 20-2). Первый этап в этой обходной последовательности реакций катализируется митохондриальной пируваткарбоксилазой. Этот биотинсодержащий фермент катализирует образование оксалоацетата из пирувата (рис. 20-3) - анаплеротическую реакцию (разд. 16.11), способную пополнять пул промежуточных продуктов цикла лимонной кислоты
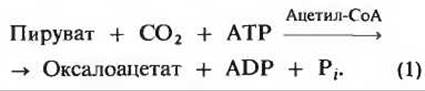
Пируваткарбоксилаза - регуляторный фермент; в отсутствие ацетил-СоА, который служит для нее положительным регулятором, она почти полностью лишена активности.
Оксалоацетат, образующийся в митохондриях из пирувата. обратимо восстанавливается за счет NADH с образованием малата под действием митохондриальной малатдегидрогеназы (разд. 16.5,3)
![]()
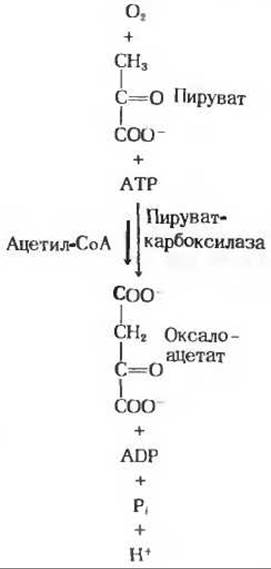
Рис. 20-3. Карбоксилирование пирувата, при водящее к образованию оксалоацетага. Включившаяся в состав оксалоацетата СО2 снова отщепляется в одной из последующих реакций (рис. 20-4).
Малат покидает митохондрии при участии специальной дикарбоксилатной транспортной системы, находящейся во внутренней митохондриальной мембране (разд. 17.19), и поступает в цитозоль. Здесь он реокисляется под действием цитозольной формы NAD-зависимой малатдегидрогеназы с образованием оксалоацетата, теперь уже внемитохондриального:
![]()
На образовавшийся таким путем оксалоацетат действует фосфоенолпируват-карбоксикиназа (разд. 16.11). Продуктом этой Mg2+-зависимой реакции, в которой донором фосфата служит гуанозинтриосфат (GTP), является фосфоенолпируват (рис. 20-4).
![]()
При внутриклеточных условиях эта реакция обратима. В клетках печени крысы фосфоенолпируват-карбоксикиназа найдена только в цитозоле, но в печени некоторых других видов этот фермент обнаруживается и в цитозоле, и в митохондриях.
Теперь мы можем написать суммарное уравнение для этих обходных реакций, посредством которых из пирувата образуется фосфоенолпируват, т.е. для реакций (1)—(4)

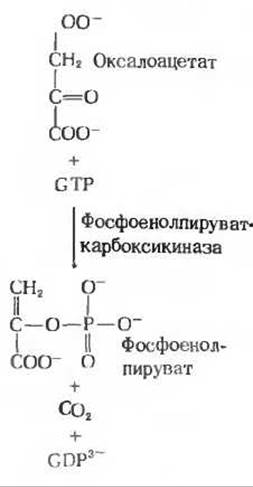
Рис. 20-4. Превращение оксалоацетата в фосфоенолпируват. Фиксированная в пируваткарбоксилазной реакции СО2 (рис. 20-3) теперь снова отщепляется в виде СО2-
Мы видим, что на фосфорилирование одной молекулы пирувата до фосфоенолпирувата (на этот процесс в стандартных условиях расход энергии составляет 14,8 ккал/моль) затрачивается энергия двух высокоэнергетических фосфатных групп (одной от АТР и одной от GTP), каждая из которых характеризуется величиной ∆G0' гидролиза -7,3 ккал/моль. Между тем, когда фосфоенолпируват превращается в пируват в процессе гликолиза, из ADP синтезируется только одна молекула АТР. Хотя изменение стандартной свободной энергии ∆G0' суммарной реакции образования фосфоенолпирувата составляет +0,2 ккал/моль, истинное изменение свободной энергии ∆G' в условиях клетки выражается очень большой отрицательной величиной, около — 6,0 ккал; ясно, таким образом, что эта реакция практически необратима.