Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Метаболизм. Общий обзор
Промежуточные стадии метаболизма можно выявлять с помощью мутантных организмов
В горой важный подход к выяснению метаболических путей связан с изучением мутантных организмов, не способных синтезировать данный фермент в активной форме. Такой дефект, если только он не является летальным, может проявиться в том, что у мутанта будет накапливаться и выводиться из организма субстрат дефектного фермента. Некоторые этапы обмена аминокислот удалось, например, выяснить, исследуя у людей врожденные нарушения обмена, при которых в организме не вырабатывается определенный фермент (рис. 13-18). У человека такие генетические нарушения встречаются сравнительно редко и вследствие этого не могут служить объектом систематического изучения. Однако у микроорганизмов их можно вызвать искусственно, воздействуя на клетки различными мутагенными агентами (рентгеновскими лучами или определенными химическими соединениями), способными изменять структуру определенных генов в их ДНК. Полученные таким путем мутантные микроорганизмы, утратившие способность синтезировать тот или иной фермент, могут служить прекрасным орудием для изучения метаболизма.
Познакомимся теперь с тем, как используются такие мутанты. Нормальные, т.е. немутантные клетки хлебной плесени Neurospora crassa (рис. 13-19), могут расти на простой среде, содержащей в качестве единственного источника углерода глюкозу и единственного источника азота - аммиак. Однако если споры этого гриба подвергнуть рентгеновскому облучению, то среди возникших мутантов обнаружатся и такие, которые уже не способны расти на этой простой среде, но вполне нормально растут на среде, содержащей определенные метаболиты. Так, например, некоторые мутанты Neurospora нормально развиваются на среде, содержащей аминокислоту аргинин, которая немутантным клеткам не требуется. Ясно, что у таких мутантов не активен или вообще не вырабатывается какой-то из ферментов, участвующих в синтезе аргинина из аммиака. Из-за отсутствия аргинина мутантные клетки не в состоянии образовать белки, в состав которых входит аргинин, и потому не растут. Однако они будут нормально расти, если мы добавим к питательной среде помимо глюкозы и аммиака еще и аргинин. Такие мутанты с нарушением какого-либо биосинтетического пути, рост которых можно восстановить, обеспечив их нормальным продуктом данного процесса, называют ауксотрофными мутантами (от греч. «aukso»-повышать; имеются в виду повышенные пищевые потребности этих мутантов).
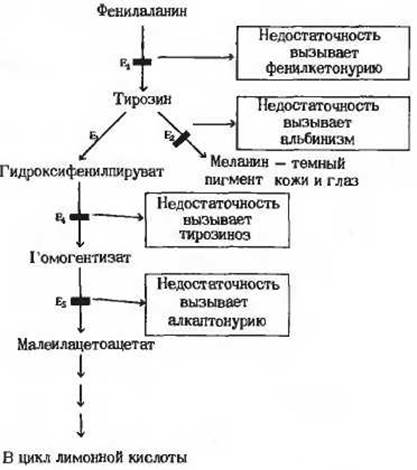
Рис. 13-18 Случаи генетического нарушения обмена аминокислоты фенилаланина, наблюдавшиеся у человека. Каждый из таких случаев связан с выпадением функции одного-единственного гена. Изучение этих нарушений дало возможность установить природу промежуточных продуктов обмена фенилаланина.
Рис. 13-19. Вегетативная форма, или мипелий, хлебной плесени Neurospora crassa. У этого организма легко получить мутантные штаммы, которые оказались весьма полезным инструментом при изучении некоторых метаболических путей. На основании экспериментов с мутантами Neurospora была сформулирована гипотеза «один ген-один фермент».
Не все мутанты Neurospora, утратившие способность синтезировать аргинин, одинаковы; они различаются в зависимости от того, какая стадия биосинтеза аргинина у них нарушена (рис. 13-20). Набор различных мутантов, нуждающихся в аргинине, можно использовать для выявления промежуточных стадий в последовательности ферментативных реакций, из которых слагается синтез этой аминокислоты. Если мутант I (рис. 13-20) выращивать на среде с очень небольшим (лимитирующим) количеством аргинина, то клетки растут до тех пор, пока весь имеющийся аргинин не будет израсходован на синтез белка. Одновременно в культуральной среде накапливается предшественник D, который не может превратиться в аргинин, поскольку у мутанта I блокирована именно та ферментативная реакция, в результате которой происходит это превращение. Удалим теперь фильтрованием из культуральной среды клетки мутанта I и поместим в нее клетки мутанта ІІ, тоже нуждающегося в аргинине, но уже по другой причине - из-за неспособности синтезировать промежуточный продукт D. Культуральная среда от мутанта I будет, очевидно, поддерживать рост мутанта II, потому что в ней присутствует предшественник D. Однако отфильтрованная культуральная среда мутанта II не должна поддерживать рост мутанта I. Мы можем поэтому определить предшественник, накапливающийся у мутанта I, по скорости роста мутанта II. Именно этим путем удалось в конечном счете идентифицировать все четыре предшественника аргинина-А, В, С и D. В подобных экспериментах по перекрестному питанию с использованием ауксотрофных мутантов Neurospora crassa или Escherichia coli были выяснены пути биосинтеза многих аминокислот.
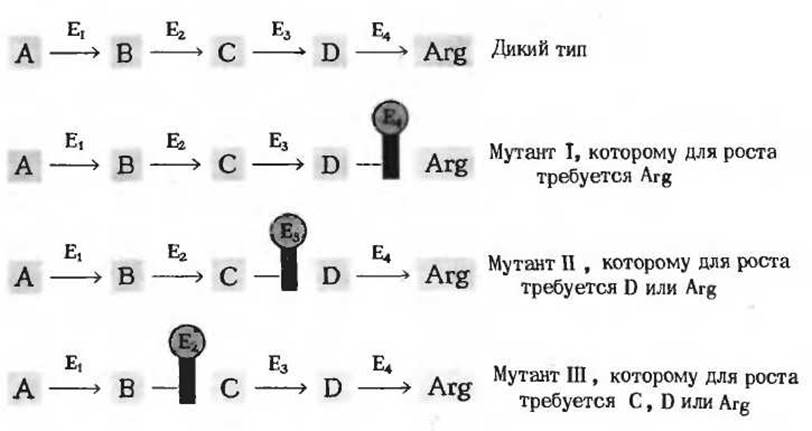
Рис. 13-20. Ауксотрофные мутанты Neurospora crassa, утратившие в результате мутации способность синтезировать один из ферментов (отмечен на рисунке красной полосой), участвующих в биосинтезе аргинина (Arg) из предшественника А. Вещества В, С и D играют роль промежуточных продуктов в этом превращении. У мутанта I отсутствует фермент Е4, но его можно выращивать на среде, обогащенной аргинином. В этих условиях в культуральной среде будет накапливаться промежуточный продукт D. У мутанта II отсутствует фермент Е3, однако активность Е4 у него сохраняется. Поэтому он может расти, если добавить к среде либо аргинин, либо промежуточный продукт D, вырабатываемый мутантом I. Аналогичным образом мутант III, лишенный фермента Е2, растет на среде, содержащей С, D или аргинин, поскольку он может превращать в аргинин промежуточные продукты С или D.