Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Биосинтез липидов
Холестерол и другие стероиды также синтезируются из двухуглеродных предшественников
Холестерол - это не только важный компонент некоторых клеточных мембран (разд. 12.10) и липопротеинов плазмы крови (разд. 12.8), но и предшественник многих других биологически важных стероидов - желчных кислот и различных стероидных гормонов. Так же как и длинноцепочечные жирные кислоты, холестерол синтезируется из ацетил-СоА, однако в этом случае ацетильные группы соединяются друг с другом иначе. Этот вывод был сделан на основе результатов экспериментов, в которых животным скармливали ацетат, меченный радиоактивным углеродом (14С) по метальной группе, и ацетат, меченный тем же изотопом по карбоксильной группе. Из тканей животных, в пищу которых добавляли меченые молекулы этих двух типов, был выделен меченый холестерол. Путем его последовательного расщепления при помощи известных химических реакций были получены характерные продукты. Определение радиоактивности этих продуктов позволило установить места локализации в молекуле холестерола атомов углерода, происходящих из метальных и карбоксильных групп. Результаты этих пионерских экспериментов, проведенных Конрадом Блоком, Робертом Вудвордом и другими исследователями, показаны на рис. 21-23. Полученная информация послужила ключом для выяснения последовательности ферментативных реакций в процессе биосинтеза холестерола, протекающего в несколько этапов (рис. 21-24).
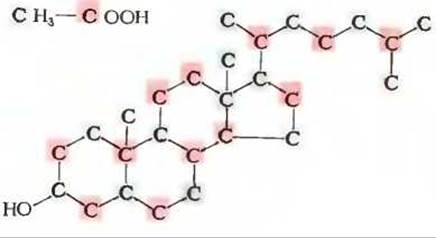
Рис. 21-23. Происхождение атомов углерода в молекуле холестерола, установленное в экспериментах с использованием ацетата, меченного радиоактивным углеродом по метильной (выделен серым цветом) и карбоксильной (выделен красным цветом) группам.
На первом этапе в результате трех приведенных ниже реакций образуется мевалоновая кислота (рис. 21-25):

В ходе последующих реакций, составляющих второй этап биосинтеза холестерола, к мевалонату присоединяются три фосфатные группы, после чего фосфорилированный мевалонат теряет карбоксильную группу и два атома водорода; в результате получается ∆3-изопентенилпирофосфат (рис. 21-26) - активированная форма изопреновой единицы (разд. 10.13). Шесть изопентенильных групп затем объединяются, теряя свои пирофосфатные группы, и образуют углеводород сквален (рис, 21-27), состоящий из 30 атомов углерода, 24 из которых соединены в цепочку, а остальные 6 входят в состав метальных боковых групп. Впервые сквален был выделен из печени акул (рода Squalus).
На третьем этапе биосинтеза холестерола происходит серия сложных ферментативных реакций, в результате которых линейная молекула схвалена превращается в циклическое соединение ланостерол, содержащее четыре типичных для стероидов конденсированных кольца (рис. 21-24). В ходе четвертой (заключительной) серии реакций ланостерол превращает я в холестерол. За расшифровку этого необычного биосинтетического пути, наиболее сложного из всех известных, американец Конрад Блок, немец Феодор Линен и англичанин Джон Корнфорт были в 1961 г. удостоены Нобелевской премии.
Регуляция биосинтеза холестерола - это также очень сложный процесс. Лимитирующей стадией служит реакция на раннем этапе биосинтеза холестерола - превращение гидроксиметилглутарил-СоА в мевалонат (рис. 21-25). Эту реакцию катализирует сложный регуляторный фермент - гидроксиметилглутарил-СоА - редуктаза, активность которого в зависимости от условий может меняться на два порядка. Этот фермент ингибируется конечным продуктом данного биосинтетического пути - холестеролом, а также мевалонатом. Гидроксиметилглутарил-СоА—редуктаза локализуется в эндоплазматическом ретикулуме; она может находиться как в фосфорилированном (неактивном), так и в нефосфорилированном (активном) состоянии. Биосинтез холестерола регулируется также концентрацией специфического белка - переносчика стеролов; этот белок связывает нерастворимые в воде промежуточные продукты биосинтеза и таким образом делает их более доступными для последующих ферментативных реакций. Скорость биосинтеза холестерола зависит не только от содержания в тканях холестерола и других стероидов; она меняется также при голодании, в зависимости от режима питания и при образовании злокачественных опухолей. Биосинтез холестерола ингибируется специфическими холестеролсодержащими липопротеинами плазмы при их связывании с соответствующими рецепторами на поверхности клеток.
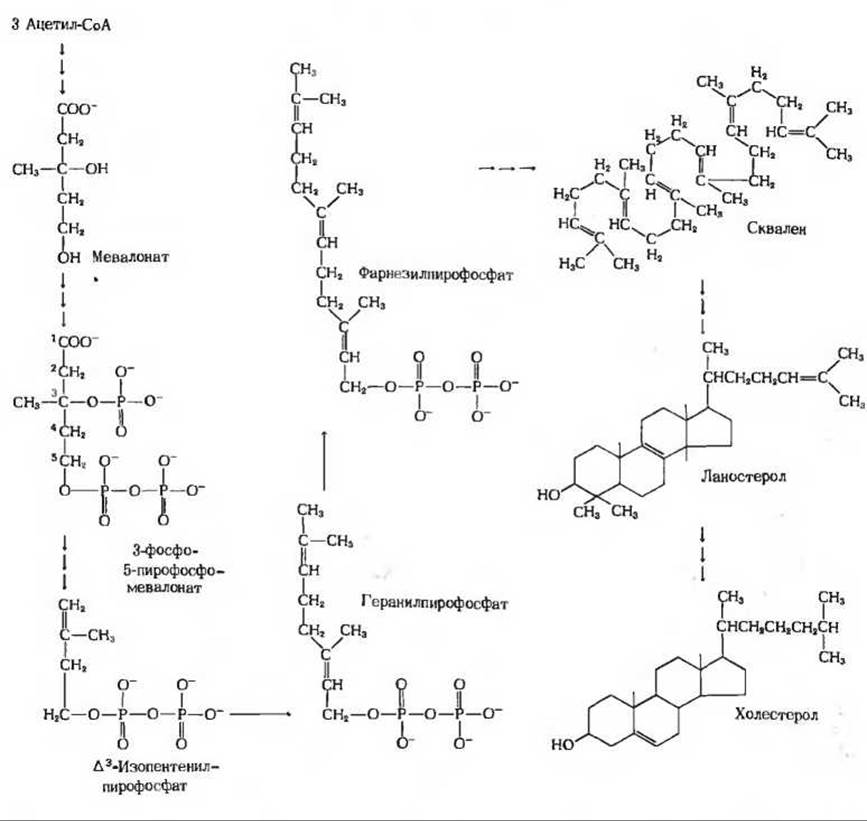
Рис. 21-24. Этапы биосинтеза холестерола. Из трех объединившихся молекул ацетил-СоА образуется мевалонат, фосфорилирование которого приводит к образованию 3-фосфо-5-пирофосфомевалоната. При отщеплении от него СО2 и фосфата получается ∆3-изопентенилпирофосфат. В результате последовательного объединения шести молекул ∆3-изопентенилпирофосфата происходит формирование линейного углеводорода сквалена, который затем циклизуется, образуя ланостерол, превращающийся в холестерол.
Нарушение регуляции биосинтеза холестерола - один из факторов, влияющих на патологический процесс атерогенеза, в ходе которого в стенках артерий и артериол образуются «бляшки», богатые холестеролом и другими липидами. Образование таких «бляшек» может приводить к нарушению кровоснабжения различных органов, чаще всего от возникающего при этом недостатка кислорода страдают сердце и мозг (гл. 26).

Рис. 21-25. Образование мевалоната из ацетил-СоА. Происходящие из ацетата 1-й и 2-й атомы углерода в молекуле мевалоната выделены красным цветом.

Рис. 21-26. Превращение мевалоната в ∆3-изопентенилпирофосфат - активированную форму изопреновой единицы. Шесть таких единиц, объединившись, образуют сквален.
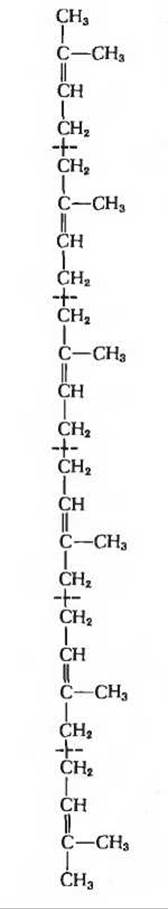
Рис. 21-27. Сквален, 30-углеродный изопреноидный углеводород, предшественник ланостерола и холестерола. Изопреновые единицы разделены цветными пунктирными линиями.