Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Биосинтез липидов
Краткое содержание главы
Длинноцепочечные насыщенные жирные кислоты синтезируются из ацетил-СоА цитоплазматическим комплексом ферментов, к числу которых относится и ацилпереносящий белок (АПБ), содержащий в качестве простетической группы фосфопантетеин. В молекуле 3-кетоацил-АПБ—синтазы имеются две существенные для ее каталитической активности —SH-группы, одна из которых принадлежит фосфопантетеину (Фп), а другая - остатку цистеина (Cys). АПБ функционирует как переносчик промежуточных продуктов синтеза жирных кислот. Образующийся из ацетил-СоА ацетил-S-Cys-AПБ реагирует с малонил-S- АПБ, образовавшимся из малонил-СоА, в результате чего получается ацетоацетил-S-АПБ и выделяется СO2. Вслед за этим происходит восстановление ацето-ацетил-S-АПБ в D-3-гидроксипроизводное, которое затем гидратируется до транс-∆2-бутеноил-S-АПБ; двойная связь в ацильной группе этого соединения восстанавливается и насыщается за счет NADPH с образованием бутирил-S-АПБ. Далее к карбоксильному концу растущей цепи жирной кислоты последовательно присоединяются еще шесть молекул малонил-S-АПБ и в результате получается пальмитоил-S—Cys-АПБ — конечный продукт реакций, катализируемых синтазным комплексом для образования жирных кислот. Затем происходит гидролитическое отщепление свободной пальмитиновой кислоты. При удлинении пальмитиновой кислоты возникает 18-углеродная стеариновая кислота. Под действием оксигеназ со смешанной функцией в молекулах пальмитиновой и стеариновой кислот возникают двойные связи, что приводит к образованию соответственно пальмитолеиновой и олеиновой кислот. Млекопитающие не способны синтезировать линолевую кислоту и должны получать ее с пищей растительного происхождения; экзогенную линолевую кислоту они могут превращать в арахидоновую кислоту, которая служит предшественником простагландинов.
Триацилглицеролы синтезируются путем взаимодействия двух молекул СоА-производных жирных кислот с глицерол-3-фосфатом; при этом образуется фосфатидная кислота, которая затем дефосфорилируется до диацилглицерола. Последний ацилируется третьей молекулой СоА-производного жирной кислоты с образованием триацилглицерола. Диацилглицеролы являются также главными предшественниками фосфоглицеролов. Голова молекулы фосфатидилэтанол-амина сначала формируется в виде цитидиндифосфатэтаноламина путем взаимодействия цитидинтрифосфата (СТР) с фосфоэтаноламином. Далее фосфоэтаноламинная группа переносится на диацилглицерол, что приводит к образованию фосфатидилэтаноламина. Фосфатидилхолин образуется путем метилирования фосфатидилэтаноламина или при взаимодействии диацилглицерола с цитидиндифосфатхолином. Холестерол также синтезируется из ацетил-СоА в результате очень сложной последовательности реакций, в которых образуются важные промежуточные соединения - гидроксиметилглутарил-СоА, мевалонат и линейный углеводород сквален. Последний циклизуется с образованием циклической стероидной системы конденсированных колец с боковой цепью. Биосинтез холестерола ингибируется поступающим с пищей холестеролом.
ЛИТЕРАТУРА
Биосинтез жирных кислот Cunningham Е.В. Biochemistry: Mechanisms of Metabolism, McGraw-Hill, New York, 1978. В гл. 12 приведено много дополнительных сведений, касающихся энзимологии и механизма синтеза жирных кислот.
Jeffcoat R. The Biosynthesis of Unsaturated Fatty Acids and Its Control in Mammalian Liver, Essays Biochem., 15, 1-36 (1979).
Биосинтез липидов
Snyder F. (ed.). Lipid Metabolism in Mammals, vols. 1 and 2, Plenum, New York, 1977.
Обмен холестерола
Bloch K.S. The Biological Synthesis of Cholesterol, Science, 150, 19-28 (1965).
Brown M. S., Goldstein J. L. Receptor-Mediated Control of Cholesterol Metabolism, Science, 191, 150-154 (1976).
Общие вопросы
Clarke M. R. The Head of the Sperm Whale, Sci. Am., 240, 128-141, January 1979. Более подробные сведения о биологической функции спермацета у кашалотов.
Nelson R. A. Protein and Fat Metabolism in Hibernating Bears, Fed. Proc., 39, 2955-2958 (1980).
Генетические нарушения
липидного обмена
Allison А. С. Lysosomes. In: J. J. Head (ed.). Carlina Biology Readers, Carolina Biological Supply Company, Burlington, N. C, 1977.
Brown M. S., Goldstein J. L. Familial Hypercholesterolemia: Defective Binding of Lipoproteins to Cultured Fibroblasts Associated with Impaired Regulation of 3-Hydroxy-3-methylglutaryl Coenzyme A Reductase Activity, Proc. Natl. Acad. Sci. USA, 71, 788-792 (1974).
Dietschy J. M, Gotto A. M, Jr., Ontko J. A. (eds.). Disturbances in Lipid and Lipoprotein Metabolism, American Physiological Society, 1978.
Stanbury J. B., Wyngaarden J. B., Fredrickson D. S. (eds.). The Metabolic Besis of Inherited Disease, 4th ed, McGraw-Hill, New York, 1978.
Вопросы и задачи
1. Роль двуокиси углерода в синтезе жирных кислот. Двуокись углерода - обязательный участник биосинтеза жирных кислот. В чем заключается специфическая роль СО2? Будет ли пальмитат, образовавшийся при инкубации растворимой фракции печени с 14СO2 и другими компонентами, необходимыми для биосинтеза жирных кислот, содержать 14С? Обоснуйте ваш ответ.
2. Путь углерода в синтезе жирных кислот. Опираясь на ваши знания о биосинтезе жирных кислот, объясните следующие экспериментальные данные:
а) Добавление равномерно меченного 14С-ацетил-СоА к растворимой фракции печени приводит к образованию равномерно меченного 14С-пальмитата.
б) Вместе с тем добавление к растворимой фракции печени следовых количеств равномерно меченного 14С-ацетил-СоА в присутствии избытка малонил-СоА приводит к образованию пальмитата, содержащего 14С только в положениях 15 и 16.
3. Суммарное уравнение синтеза жирных кислот. Напишите суммарное уравнение биосинтеза пальмитиновой кислоты в печени крысы, начиная с митохондриального ацетил-СоА и цитозольных NADPH, АТР и СO2.
4. Путь водорода в синтезе жирных кислот. Возьмем препарат, содержащий все ферменты и кофакторы, необходимые для биосинтеза жирной кислоты из ацетил-СоА и малонил-СоА.
а) Сколько атомов дейтерия (тяжелого изотопа водорода) включится в каждую молекулу пальмитиновой кислоты, если в качестве субстратов использовать меченный дейтерием ацетил-СоА
![]()
и избыток немеченого малонил-СоА? В каких положениях они будут находиться? Ответ объясните, б) Сколько атомов дейтерия включится в каждую молекулу пальмитата, если в качестве субстратов использовать немеченый ацетил-СоА и меченный дейтерием малонил-СоА
![]()
В каких положениях они будут находиться? Дайте объяснение.
5. Образование NADPH, необходимого для биосинтеза жирных кислот. Поскольку внутренняя мембрана митохондрий непроницаема для ацетил-СоА, ацетильные группы попадают в цитозоль посредством челночного переноса (см. схему на рис. 21-3). В цитозоле содержится NADP-зависимая малатдегидрогеназа, катализирующая реакцию
![]()
Считая, что внутренняя мембрана митохондрий проницаема для пирувата (а также для цитрата и малата), предложите челночную схему образования NADPH в цитозоле при помощи NADP-зависимой малатдегидрогеназы и других ферментов, о которых известно, что они локализованы в митохондриях и цитозоле. Напишите суммарную реакцию переноса ацетильных групп из митохондрий в цитозоль.
6. Активность ацетил-СоА—карбоксилазы изменяется под действием модуляторов. Ацетил-СоА—карбоксилаза является ключевым регуляторным пунктом биосинтеза жирных кислот. Некоторые характеристики этого фермента приведены ниже.
а) Добавление цитрата или изоцитрата повышает Vmах фермента более чем в 10 раз.
б) Фермент существует в двух резко различающихся по активности формах, которые могут превращаться друг в друга.
![]()
Цитрат и изоцитрат связываются предпочтительно с нитевидной формой фермента, а пальмитоил-СоА-с протомерной.
Объясните, как эти свойства связаны с регуляторной ролью ацетил-СоА—карбоксилазы в биосинтезе жирных кислот.
7. Сколько «стоит» синтез триацилглицеролов. Исходя из суммарного уравнения биосинтеза трипальмитина из глицерола и пальмитиновой кислоты, подсчитайте, сколько молекул АТР требуется для образования одной молекулы грипальмитина.
8. Энергетические затраты в ходе синтеза фосфатидилхолина. Напишите последовательные этапы и суммарное уравнение для биосинтеза фосфатидилхолина по «спасательному» пути из олеиновой и пальмитиновой кислот, дигидроксиацетонфосфата и холина. Сколько молекул АТР потребуется для синтеза фосфатидилхолина этим способом, если начинать его с олеиновой кислоты, пальмитиновой кислоты и дигидроксиацетонфосфата?
9. Лечение гиперхолестеринемии. Растения не синтезируют холестерол, а вырабатывают другие стеролы, называемые фитостеролами. Строение одного из них - ß-ситостерола - здесь показано. Когда больные с гиперхолестеринемией получают с пищей ß-ситостерол, уровень холестерола в плазме у них снижается, что должно уменьшать вероятность заболевания атеросклерозом. Предложите возможные механизмы действия ß-ситостерола.
10. Взаимосвязь метаболизма аминокислот с метаболизмом жирных кислот. Крысе ввели препарат 3-14С-аланина
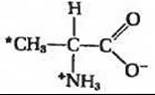
Через час после этого животное забили и из печени экстрагировали липиды. Полученный при экстракции пальмитат содержал 14С. Как это можно объяснить? В каком месте молекулы пальмитата находится 14С? Может ли аланин служить предшественником в реальном синтезе пальмитата de novo?
11. Различия между анаболическими и катаболическими путями жирных кислот.
Один оборот анаболического и катаболического циклов короткоцепочечных жирных кислот можно представить уравнением
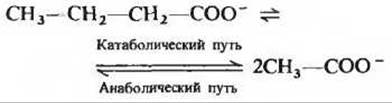
а) Сравните суммарные уравнения катаболического и анаболического путей. Являются ли они просто обратными реакциями по отношению друг к другу? В чем их различия?
б) Какие специфические факторы позволяют этим путям идти независимо друг от друга?