Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Фотосинтез
Фотосинтетическое фосфорилирование сходно с окислительным фосфорилированием
Фотосинтетический перенос электронов и фотофосфорилирование в хлоропластах во многом сходны с переносом электронов и окислительным фосфорилированием в митохондриях. Это сходство проявляется в следующем: 1) реакционные центры, переносчики электронов и ферменты, участвующие в образовании АТР, находятся в мембране тилакоидов; 2) необходимым условием фотофосфорилирования является целостность тилакоидных мембран; 3) тилакоидная мембрана непроницаема для ионов Н+; 4) фотофосфорилирование можно разобщить с переносом электронов при помощи реагентов, способных стимулировать прохождение ионов Н+ через тилакоидную мембрану; 5) фотофосфорилирование можно блокировать олигомицином и другими аналогичными агентами, подавляющими в митохондриях синтез АТР из ADP и фосфата, катализируемый АТР-синтетазой (разд. 17.14); 6) синтез АТР осуществляется «грибовидными» ферментными молекулами, находящимися на наружной поверхности тилакоидной мембраны; по своей структуре и функции они очень напоминают F1-ATPaзy митохондрий, вследствие чего их часто обозначают символом CF1 (от англ. chloroplast - хлоропласт).
Подобно внутренней митохондриальной мембране (рис. 17-2), тилакоидная мембрана асимметрична по-своему молекулярному строению (рис. 23-15). Молекулы переносчиков в цепи переноса электронов, ведущей от фотосистемы II к фотосистеме I, ориентированы в тилакоидной мембране таким образом, что перенос электронов создает реальный поток ионов Н+ через тилакоидную мембрану, направленный снаружи внутрь. Под влиянием индуцированного светом потока электронов возникает, следовательно, трансмембранный градиент концентрации ионов Н+, так что с внутренней стороны тилакоидных пузырьков среда становится более кислой, чем с наружной. Все эти свойства согласуются с хемиосмотической гипотезой, предложенной первоначально для объяснения окислительного фосфорилирования, а позднее распространенной и на фотосинтетическое фосфорилирование. Схема на рис. 23-16 изображает поток ионов Н+, движимых световой энергией; ионы Н+ поступают из стромы внутрь тилакоида и вновь возвращаются наружу через молекулы АТР-синтетазы.
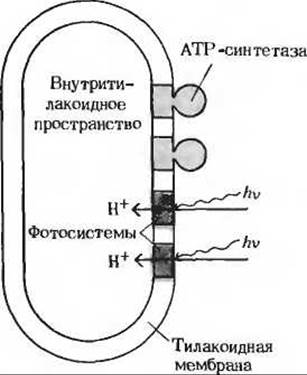
Рис. 23-15. Внутренняя и наружная поверхности тилакоидной мембраны различны. Фотосистемы и цепь переноса электронов ориентированы таким образом, что они накачивают ионы Н+ внутрь тилакоида. АТР-синтетазные «головки» (CF1) располагаются на наружной поверхности мембраны.
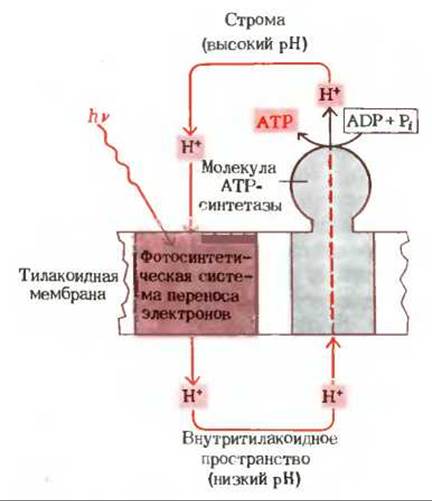
Рис. 23-16. Фотосинтетический Н+-цикл. Фотосинтетический перенос электронов вызывает перекачивание ионов Н+ из наружной среды в тилакоид, в результате чего возникает трансмембранный Н+-градиент, обращенный кислым концом внутрь. Возврат ионов Н+ наружу через ориентированную АТР-синтетазную молекулу поставляет энергию для синтеза АТР из ADP и фосфата.
В 1966 г. Андрэ Ягендорф поставил важный эксперимент, доказавший, что источником энергии для синтеза АТР действительно может служить трансмембранный градиент pH, обращенный щелочным концом наружу. Сначала он инкубировал хлоропласта в темноте в буферном растворе (pH 4), который медленно проникал во внутренний компартмент тилакоидов, снижая pH их содержимого. Затем, после добавления к этой темновой суспензии хлоропластов ADP и фосфата, он быстро доводил pH среды до 8, добавляя к ней щелочной буфер, т. е. мгновенно создавал большой трансмембранный градиент pH. Когда этот градиент начинал уменьшаться вследствие выхода ионов Н+ из гилакоидов в среду, происходило образование АТР из ADP и фосфата. Поскольку АТР синтезировался в темноте, этот эксперимент доказывал, что трансмембранный градиент pH есть высокоэнергетическое состояние, посредством которого энергия переноса электронов передается АТР-синтетазе, получающей таким образом возможность генерировать АТР, как это постулируется хемиосмотической гипотезой.