Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
ATP-Цикл и биоэнергетика клетки
Креатинфосфат в мышцах выполняет роль резервуара высокоэнергетических фосфатных групп
Среди высокоэнергетических фосфорилированных соединений имеется одно, играющее особую роль в энергетике возбудимых тканей, таких, как мышечная и нервная. Это соединение, креатинфосфат, или фосфокреатин (рис. 14-13), служит резервуаром высокоэнергетических фосфатных групп. ∆С0' гидролиза креатинфосфата (-10,3 ккал/моль) несколько превышает ∆С0' гидролиза АТР. Креатинфосфат может передавать свою фосфатную группу на ADP в реакции, катализируемой креатинкиназой:
![]()
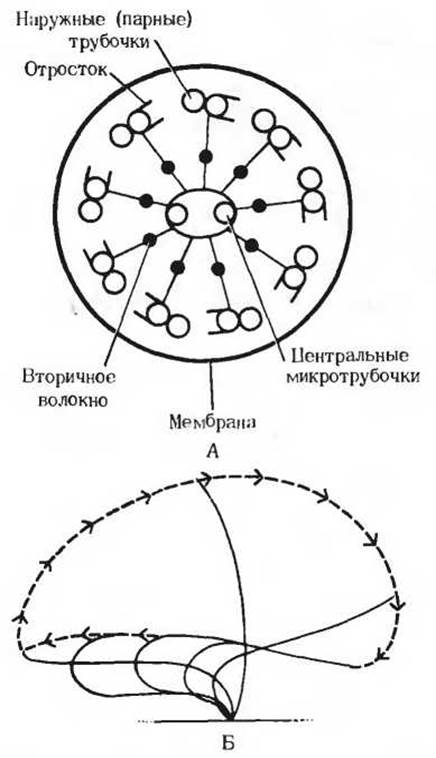
Рис. 14-12. В ресничках и жгутиках эукариотических клеток механическая сила развивается за счет использования АТР. А. Поперечный разрез реснички. Эти структуры состоят из девяти пар микротрубочек, образующих наружное кольцо, и двух одиночных центральных микротрубочек (расположение по типу «9 + 2»; разд. 2.16). Реснички окружены оболочкой, представляющей собой вырост клеточной мембраны. Энергию для характерных движений ресничек (волнообразного, скользящего или вращательного) поставляет гидролиз АТР. Эти движения осуществляются ресничками за счет скольжения или скручивания парных микротрубочек, которое весьма напоминает наблюдаемое в скелетных мышцах АТР-зависимое скольжение толстых и тонких нитей друг относительно друга. От наружных (парных) микротрубочек отходят находящиеся на равном расстоянии друг от друга отростки, или выступы, напоминающие миозиновые головки в толстых нитях мышц. Эти выступы состоят из молекул динеина - довольно крупного белка, обладающего АТРазной активностью. Катализируемый динеином гидролиз АТР поставляет энергию для механического движения - скольжения или скручивания микротрубочек. Было высказано предположение, что центральные микротрубочки регулируют скорость движения ресничек. Б. Отдельные фазы биения реснички в жабрах морского червя, у которого реснички имеют длину около 30 мкм. Эти характерные движения сообщает ресничкам АТР-зависимое скольжение трубчатых нитей друг относительно друга.
Благодаря креатинфосфату концентрация АТР в мышечных клетках поддерживается на постоянном и притом довольно высоком уровне. Особенно это существенно для скелетных мышц, работающих с перерывами, но иногда очень напряженно с большой скоростью. Всякий раз, когда часть АТР мышечной клетки расходуется на сокращение, в результате гидролиза АТР образуется ADP. Креатинфосфат при участии креатинкиназы быстро передает свою фосфатную группу молекулам ADP, и нормальный уровень АТР восстанавливается. Содержание креатинфосфата в мышцах в 3-4 раза превышает содержание АТР (табл. 14-4); поэтому в форме креатинфосфата может храниться достаточное количество фосфатных групп, полностью обеспечивающее поддержание постоянного уровня АТР в короткие периоды усиленной мышечной активности. Благодаря обратимости креатинкиназной реакции накопившийся креатин в период восстановления вновь фосфорилируется за счет АТР до креатинфосфата. Поскольку другого метаболического пути для образования и расщепления креатинфосфата не существует, это соединение хорошо приспособлено для выполнения своей функции - резервуара фосфатных групп.
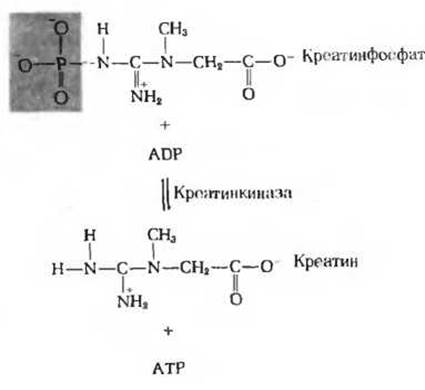
Рис. 14-13. Креатинфосфат в мышцах играет роль запасного донора высокоэнергетических фосфатных групп. Он действует как своеобразный буфер, обеспечивающий постоянство концентрации АТР.
В мышцах многих беспозвоночных роль носителя резервной формы энергии выполняет не креатинфосфат, а аргининфосфат. Соединения, служащие, подобно креатинфосфату и аргининфосфату, запасными источниками энергии, носят название фосфагенов.