Основы биохимии Том 3 - А. Ленинджер 1985
Некоторые аспекты биохимии человека
Пищеварение, транспорт питательных веществ и взаимосвязь обменных процессов
Мозг использует энергию дли передачи импульсов
Метаболизм мозга примечателен в ряде отношений. Во-первых, в качестве клеточного топлива мозг взрослых млекопитающих обычно использует только глюкозу. Во-вторых, мозгу свойствен очень интенсивный дыхательный обмен; так, у взрослого человека мозгом используется почти 20% кислорода, потребляемого организмом в состоянии покоя. Кроме того, скорость использования кислорода мозгом отличается большим постоянством и не претерпевает существенных изменений при переходе, например, от активной умственной работы ко сну. Из-за низкого содержания гликогена мозг нуждается в постоянной доставке глюкозы кровью. Если концентрация глюкозы в крови даже на короткий период времени оказывается существенно ниже определенного критического уровня, то могут возникнуть тяжелые и иногда необратимые нарушения функции мозга. Именно поэтому хирургические операции на мозге возможны только при условии, что обеспечено его бесперебойное снабжение глюкозой крови.
Хотя мозг не может прямо использовать свободные жирные кислоты или липиды крови в качестве клеточного топлива, однако он утилизирует доставляемый кровью ß-гидроксибутират, образующийся в печени из жирных кислот. Способность мозга окислять ß-гидроксибутират через промежуточное образование ацетил-СоА (разд. 18.10) приобретает особо важное значение при продолжительном голодании, когда практически весь гликоген печени оказывается израсходованным; в этих условиях мозг переходит на использование в качестве источника энергии имеющихся в организме жиров. Запасы жира в организме намного превышают запасы гликогена; при голодании запасов гликогена хватает всего лишь на несколько дней. Использование мозгом ß-гидроксибутирата во-время голодания позволяет также сохранить белки мышц, которые - через процесс глюконеогенеза - служат для мозга последним источником глюкозы при голодании.
Глюкоза используется мозгом в ходе гликолиза и в цикле лимонной кислоты; распад глюкозы обеспечивает почти весь запас АТР мозга. За счет энергии АТР нервные клетки (нейроны) поддерживают электрический потенциал на плазматической мембране и, в частности, на мембране, окружающей их длинные отростки-аксоны и дендриты, образующие “линии передач” в нервной системе. Передача нервных импульсов вдоль нейронов происходит посредством волнообразного изменения электрических свойств мембраны, т. е. так называемого потенциала действия. Na+, К+ -АТРаза плазматической мембраны (разд. 14.16) нуждается в постоянном притоке энергии АТР для накачивания ионов К+ внутрь аксонов и выведения ионов Na+ из аксонов (рис. 24-14). За счет энергии гидролиза одной молекулы АТР три иона Na+ выводятся наружу, а два иона К+ поступают внутрь аксона. Такой дисбаланс электрических зарядов служит причиной того, что Na+, К+ -АТРаза генерирует разность потенциалов на мембране аксонов, причем обычно наружная сторона мембраны заряжена положительно.
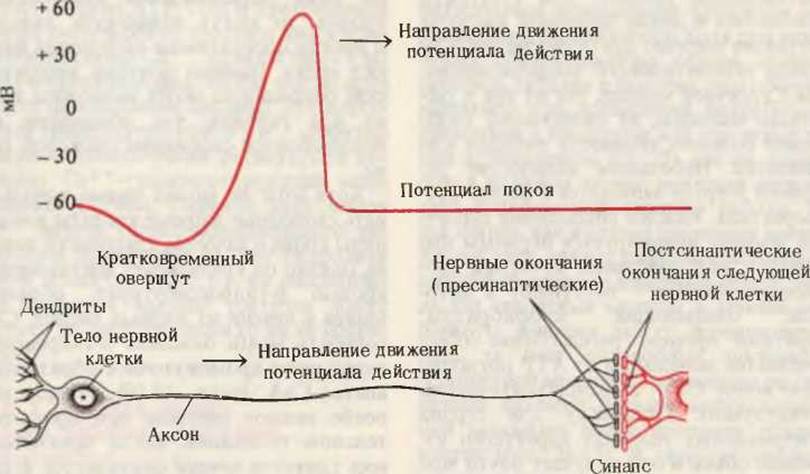
Рис. 24-14. Нейрон и потенциал действия. Поступающие на дендриты импульсы передаются вдоль аксона к следующему нейрону. Потенциал покоя обычно равен — 60 мВ (отрицательный заряд - на внутренней стороне мембраны). Изменение знака заряда происходит вследствие быстрого тока ионов Na+ из внеклеточного пространства внутрь клетки, обусловленного избирательным открытием Nа+-каналов. Восстановление потенциала покоя обеспечивается действием Na+, К+ -транспортирующей АТРазы, локализованной в мембране аксона.
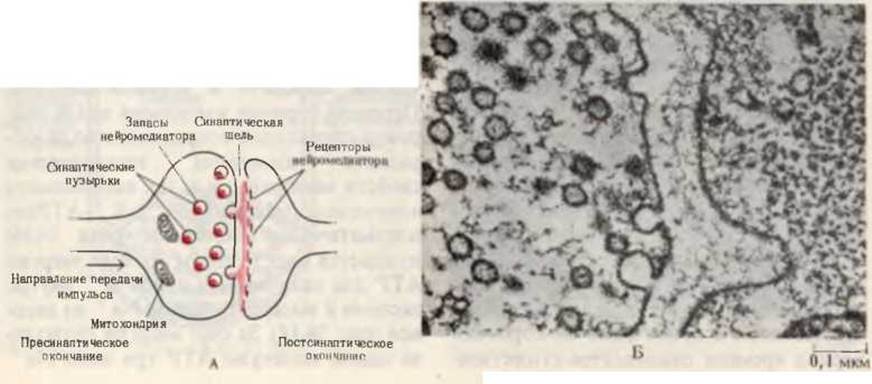
Рис. 24-15. А. Схема синапса, на которой показаны его основные функциональные компоненты. Митохондрии вырабатывают АТР, необходимый для концентрирования нейромедиатора в секреторных везикулах, а также энергию, исполняемую для обратного всасывания нейромедиатора из синаптической щели. Б. Электронная микрофотография, на которой видно высвобождение нейромедиаторов из секреторных везикул в нервно-мышечное соединение.
Большое количество АТР в мозгу используется для синтеза нейромедиаторов - веществ, которые передают импульсы с одного нейрона на другой через синапсы, т. е. через участки контакта между последовательно расположенными нервными клетками. Известно большое число как нейромедиаторов, так и веществ, ингибирующих передачу импульсов; каждое из них специфично в отношении того или иного типа нейронов либо определенных участков мозга. К числу таких веществ относятся глутамат, глутамин, аспартат, глицин и у-аминобутират. В качестве нейромедиаторов либо ингибиторов могут функционировать производные других аминокислот, а также некоторые пептиды и ацетилхолин. Нейромедиаторы накапливаются в специальных везикулах (пузырьках) в пресинаптических нервных окончаниях (рис. 24-15). В ответ на потенциал действия, возникший на мембране аксона, содержимое отдельных пузырьков изливается в синаптическую щель и связывается со специфическими рецепторными участками в чувствительном окончании постсинаптического нейрона, тем самым стимулируя его к дальнейшей передаче импульса. После стимуляции постсинаптического нейрона высвобожденный в синаптическую щель нейромедиатор должен быть либо быстро разрушен ферментами, либо реабсорбирован в пресинаптическое нервное окончание, иначе синапс не будет способен к проведению следующего импульса. Ферментативная инактивация нейромедиатора ацетилхолина осуществляется в синаптической щели с помощью ацетилхолинэстеразы, гидролизующей этот медиатор до ацетата и свободного холина (разд. 9.12).