Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Репликация и транскрипция ДНК
ДНК реплицируется полуконсервативным способом
Теперь, когда мы изучили структуру ДНК и природу хромосом и генов, посмотрим, как происходит репликация ДНК с образованием дочерних молекул и транскрипция с образованием комплементарных ей РНК.
Ферменты и прочие белки, участвующие в процессах репликации и транскрипции ДНК, относятся к числу самых замечательных из всех известных биологических катализаторов. Они способны создавать эти гигантские макромолекулы из мононуклеотидов-предшественников, используя энергию фосфатной группы, и с исключительной точностью осуществлять перенос генетической информации от матрицы к новосинтезируемой цепи. Кроме того, при работе этих ферментов должны решаться сложные механические проблемы, поскольку, прежде чем в дело вступят реплицирующие ферменты, должно произойти расплетание родительской двухцепочечной ДНК так, чтобы ферменты могли получить доступ к информации, закодированной в последовательности оснований внутри двойной спирали. Более того, в эукариотических клетках система репликации тесно связана со сложной трехмерной организацией хроматина и нуклеосом.
Ферменты транскрипции также обладают необычными свойствами. Они способны не только катализировать синтез широкого набора различных РНК, но и начинать и заканчивать свое действие в особых точках хромосомы, реагируя на разнообразные регуляторные сигналы. В результате согласованного действия ферментов на определенных этапах жизненного цикла клетки транскрибируются только определенные гены. Таким образом, ДНК- и РНК-полимеразы, как, впрочем, и другие белки, помогающие осуществлять репликацию и транскрипцию ДНК, жизненно важны для сохранения в поколениях генетической информации.
Согласно гипотезе Уотсона-Крика, каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской ДНК, причем каждая из этих молекул содержит одну неизмененную цепь родительской ДНК. Гипотеза Уотсона-Крика была проверена с помощью остроумных опытов, выполненных Мэтью Мезельсоном и Франклином Сталем в 1957 г. Основная идея этих опытов иллюстрируется схемой, приведенной на рис. 28-1. Клетки Е. coli выращивали в течение ряда поколений в среде, содержащей в качестве источника азота хлористый аммоний (NH4Cl), в котором обычный распространенный изотоп [14N] был заменен на “тяжелый” изотоп [15N]. Вследствие этого все соединения клеток, имеющие в своем составе азот, в том числе и основания ДНК, оказались сильно обогащенными изотопом [15N], Плотность ДНК, выделенной из этих клеток, была приблизительно на 1% выше плотности нормальной [14N] ДНК. Хотя это различие невелико, тем не менее смесь “тяжелой” [15N] и “легкой” [14N] ДНК удалось разделить методом центрифугирования в концентрированном растворе хлористого цезия. Хлористый цезий использовали потому, что плотность его водного раствора может быть сделана достаточно близкой к плотности ДНК. Если такой раствор СsСl долго центрифугировать в высокоскоростной ультрацентрифуге, то через некоторое время достигается состояние равновесия, при котором в пробирке образуется непрерывный градиент плотности CsCl. Благодаря сильному гравитационному полю, создаваемому центрифугой, концентрация CsCl и соответственно плотность раствора у дна пробирки оказывается более высокой, чем наверху. Препарат ДНК, растворенный в CsCl, приходит в равновесное состояние в том месте пробирки, в котором плотность ДНК равна плотности раствора CsCl. Поскольку [15N] ДНК чуть тяжелее, чем [14N] ДНК, полоса, в которой она достигает равновесия в градиенте CsCl, расположена ближе ко дну пробирки, чем полоса с [14N] ДНК (рис. 28-2).
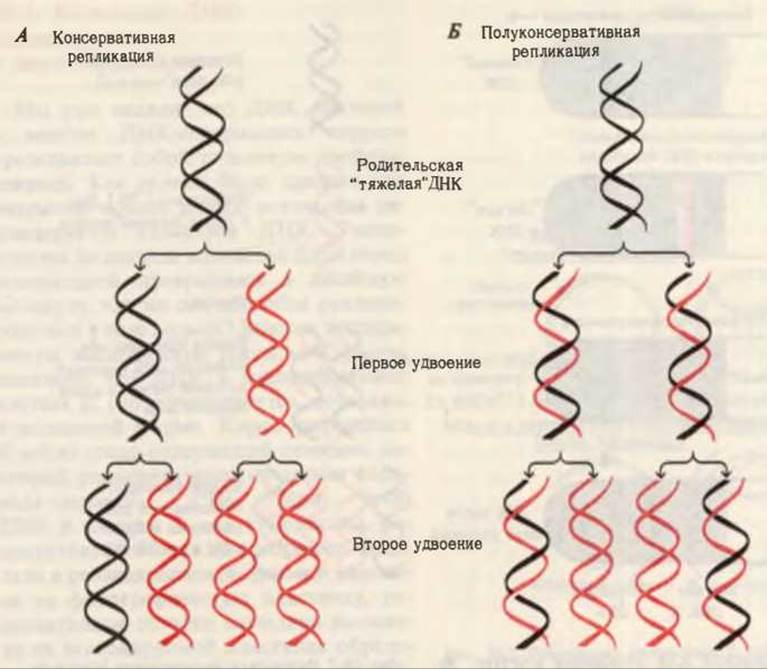
Рис. 28-1. Принцип эксперимента Мезельсона-Сталя, целью которого было выбрать один из двух возможных механизмов репликации “тяжелой” [15N] ДНК (обозначена черным цветом), когда репликация происходила в среде с “легким” изотопом азота [14N], “Легкие” цепи ДНК обозначены красным цветом. А. Консервативный механизм репликации. Если бы репликация протекала по этому механизму, то одна из двух дочерних двухцепочечных молекул содержала бы две “легкие”, а другая - две “тяжелые” цепи. Последующее удвоение дочерних молекул должно было бы привести к появлению четырех двухцепочечных молекул, одна из которых была бы “тяжелой”, а три других - “легкими”; гибридные ДНК при этом не образуются. Б. Полуконсервативный механизм репликации. При этом типе репликации каждая из двух дочерних двухцепочечных молекул ДНК должна была бы содержать одну родительскую (“тяжелую”) цепь и одну “легкую” цепь. Последующее удвоение дочерних молекул приводило бы к образованию двух гибридных и двух “легких” молекул ДНК.
Мезельсон и Сталь перенесли клетки E. coli, росшие на среде с [15N] и содержащие “тяжелые” цепи ДНК, на свежую среду, в которой находился NH4Cl с обычным изотопом [14N]. На этой среде они выращивали клетки в течение времени” необходимого для удвоения клеток. Затем из этих клеток выделяли ДНК и анализировали ее плотность с помощью описанного выше метода седиментации. В градиенте CsCl была обнаружена лишь одна полоса ДНК, плотность которой оказалась средней между плотностью нормальной “легкой” [14N] ДНК и плотностью “тяжелой” [15N] ДНК (рис. 28-2). Именно такого результата и следовало ожидать, если двухцепочечная ДНК дочерних клеток содержит одну старую 15N-цепь от родительской ДНК, и одну новую 14N-цепь (рис. 28-2).
Если выделить ДНК из клеток, которые прошли два цикла удвоения на среде с [14N], то она разделится на две полосы: одна с плотностью, соответствующей плотности нормальной легкой ДНК, а другая с плотностью гибридной ДНК, наблюдавшейся после первого удвоения клеток. На основании этих данных Мезельсон и Сталь пришли к выводу, что в строгом соответствии с гипотезой Уотсона-Крика каждый дочерний дуплекс ДНК после двух циклов удвоения клеток содержал одну родительскую и одну новообразованную цепь ДНК. Такой механизм репликации назвали полуконсервативным, поскольку в каждой дочерней ДНК сохраняется лишь одна родительская цепь (рис. 28-1 и 28-2). Полученные результаты полностью исключили консервативный способ репликации, при котором одна дочерняя ДНК должна была бы содержать обе исходные цепи, а другая состояла бы из двух новосинтезированных цепей. Опыт Мезельсона и Сталя позволил также отвергнуть так называемый дисперсивный механизм репликации, при котором каждая дочерняя цепь ДНК состоит из коротких участков как родительской, так и новообразованной ДНК, соединенных между собой случайным образом.
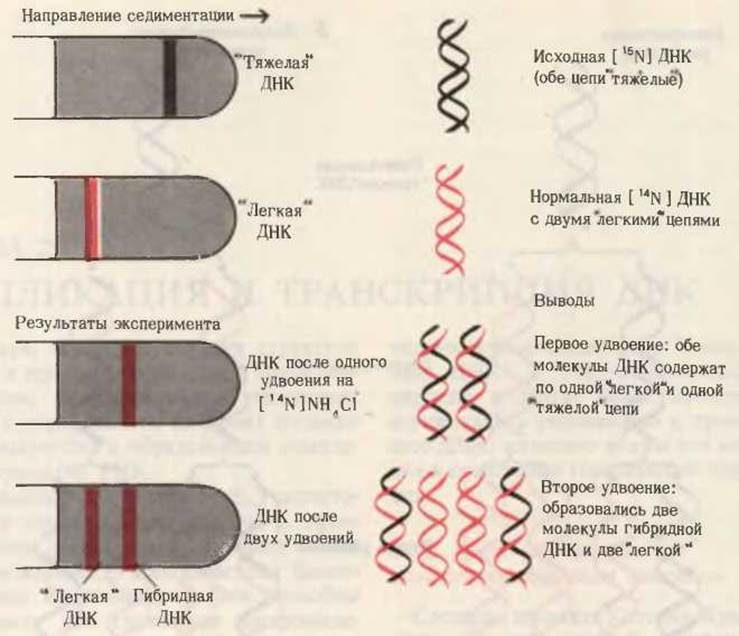
Рис. 28-2. Результаты эксперимента Мезельсо- на-Сталя. В градиенте плотности CsCl “тяжелая” [15N] ДНК достигает равновесия в полосе, которая расположена ближе ко дну пробирки, чем полоса равновесия “легкой” [14N]ДHK. Равновесное положение гибридной ДНК оказывается промежуточным. Определение плотности дочерних ДНК после первого и второго удвоений показало, что репликация ДНК осуществляется по полуконсервативному механизму.