Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Репликация и транскрипция ДНК
Для репликации необходимо физическое разделение цепей родительской двухцепочечной ДНК
Мы уже видели, что кольцевая двухцепочечная ДНК реплицируется одновременно в обоих направлениях, так что две репликативные вилки перемещаются вдоль кольцевой хромосомы навстречу друг другу. Если мы теперь вспомним, что двойная спираль ДНК представляет собой плотно скрученную структуру и что кодирующие основания находятся внутри спирали, то станет ясно: чтобы реплицирующие ДНК ферменты смогли “прочитать” нуклеотидную последовательность матрицы, цепи родительской ДНК должны быть разделены хотя бы на коротком участке. Даже если бактериальная ДНК отрицательно сверхспирализована, т. е. уже слегка раскручена, по мере движения репликативной вилки вперед она должна дополнительно расплетаться.
Раскручивание двойной спирали и удержание двух цепей на некотором расстоянии друг от друга, чтобы они могли реплицироваться, осуществляется при помощи нескольких специальных белков (рис. 28-13). Ферменты, известные под названием хеликаз (helicase, от слова helix-спираль), расплетают короткие участки ДНК, находящиеся непосредственно перед репликативной вилкой. Для раскручивания ДНК требуется энергия. На разделение каждой пары оснований расходуется энергия гидролиза двух молекул АТР до ADP и фосфата. Как только небольшой участок ДНК оказывается расплетенным, к каждой из разделившихся цепей прочно присоединяются несколько молекул ДНК-связывающего белка (ДСБ), которые препятствуют образованию комплементарных пар и обратному воссоединению цепей. Благодаря этому нуклеотидные последовательности цепей ДНК оказываются доступными для репликативной системы. ДНК-полимераза может непосредственно удлинять ведущую цепь, добавляя к ее 3'-концу новые нуклеотиды. Другие специфические белки помогают примазе получить доступ к матрице для отстающей цепи. В результате примаза получает возможность связываться с отстающей цепью и синтезировать РНК-затравки для фрагментов Оказаки. Расплетание ДНК-одна из наиболее интересных и в то же время наиболее сложных проблем репликации ДНК. Предстоит еще много сделать, прежде чем мы поймем, как живая клетка решает эту механическую и биохимическую проблему. На рис. 28-13 указаны основные белки, необходимые для репликации ДНК.
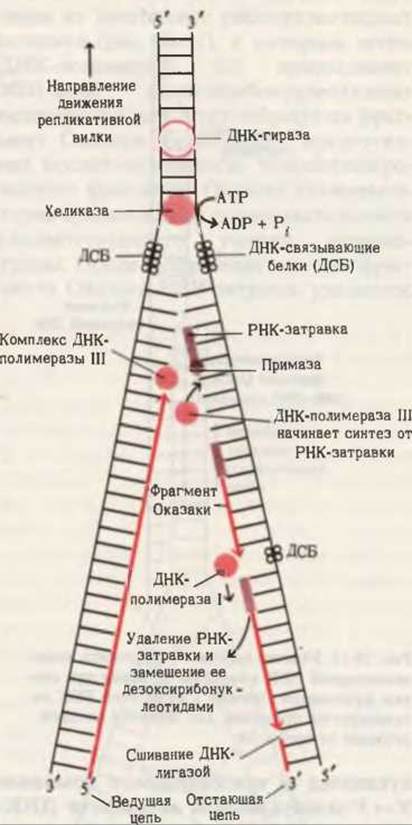
Рис. 28-13. Схематическое изображение основных этапов репликации ДНК. Существует некоторая неопределенность относительно точного места действия ДНК-гиразы.
Быстрое раскручивание цепей родительской ДНК в процессе репликации (4500 об/мин) порождает еще одну проблему, которая состоит в том, что при отсутствии специального шарнирного устройства вся хромосома, расположенная впереди репликативной вилки, должна вращаться с такой же скоростью. Предполагают, что избежать этого помогает клетке шарнир в ДНК (возможно, прямо перед репликативной вилкой), благодаря которому вращаться с большой скоростью приходится только короткому участку ДНК. Это может быть достигнуто за счет кратковременного разрыва одной из цепей ДНК, который очень быстро и точно восстанавливается после одного или нескольких оборотов. Кратковременные разрывы и воссоединения осуществляются ферментами, известными под названием топоизомераз. У прокариот топоизомераза называется ДНК-гиразой (от англ. gyration - вращение). Этот фермент не только позволяет ДНК вращаться, но и активно закручивает ее в направлении, благоприятствующем расплетанию цепей матрицы в районе репликативной вилки. Таким образом, гираза помогает хеликазе раскручивать ДНК для ее репликации. Закручивание ДНК с помощью гиразы и сопряженный с этим процессом гидролиз АТР до ADP и Рi, обусловливают сверхспиральное состояние хромосомы. Благодаря гиразе все кольцевые ДНК бактериальных клеток поддерживаются в сверхспиральной форме (рис. 28-14).
По мере того как репликативная система ликвидирует разрывы в отстающей цепи, она движется вдоль реплицирующейся ДНК. Две новые цепи соединяются со своими комплементарными цепями-матрицами автоматически, образуя две дочерние двойные спирали, каждая из которых содержит одну родительскую и одну новосинтезированную цепь. Для образования новых спиралей не требуется ни затрат энергии, ни участия какого-либо “закручивающего” фермента.
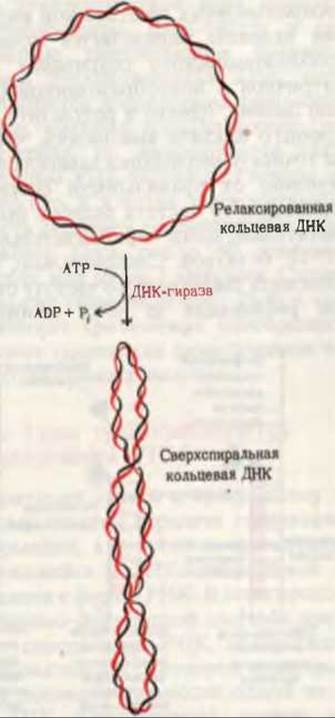
Рис. 28-14. Образование отрицательных сверхвитков под действием ДНК-гиразы. Необходимая для этого процесса энергия освобождается в результате гидролиза АТР. ДНК-гираза делает в одной из цепей ДНК разрыв. Через открывшуюся брешь выходит интактная цепь, после чего концы разорванной цепи сшиваются тем же ферментом.