Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Репликация и транскрипция ДНК
Транскрипты РНК претерпевают дальнейшие превращения
Транскрипты РНК, синтезированные при помощи РНК-полимеразы, обычно претерпевают дальнейшие ферментативные превращения, называемые посттранскрипционным процессингом, и только после этого они обретают свою функциональную активность. рРНК и тРНК синтезируются в виде более длинных предшественников, которые затем модифицируются и расщепляются с образованием конечных продуктов. Транскрипты эукариотических мРНК также подвергаются процессингу, тогда как у прокариотических мРНК он отсутствует.
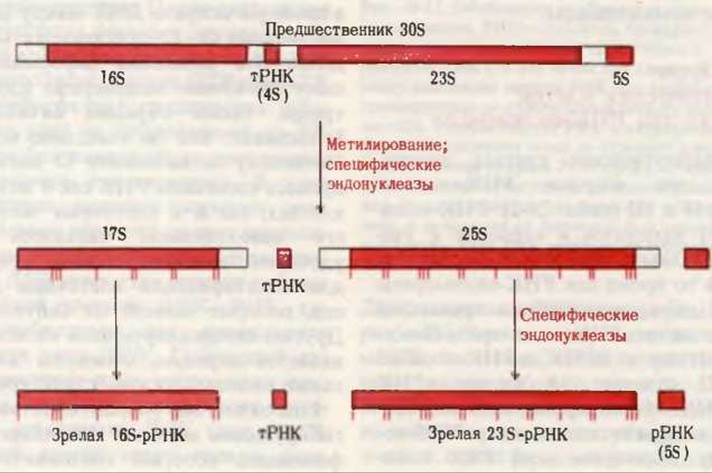
Рис. 28-19. Процессинг (“созревание”) транскрипта рРНК у прокариот. Зрелые молекулы 16S- и 23S-pPHK образуются из более длинного предшественника - 30S-PHK в результате действия специфических нуклеаз. Перед расщеплением 30S-PHK метилируется по определенным основаниям (отмечены красными вертикальными черточками). Из средней части предшественника образуется одна молекула тРНК.
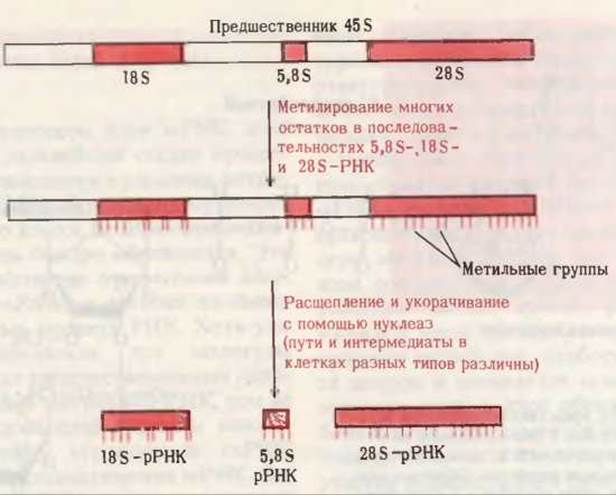
Рис. 28-20. Процессинг транскриптов эукариотических рРНК. Метилирование, представляющее собой первый этап процессинга, осуществляется по 2’-гидроксильным группам рибозных остатков в тех участках предшественника, которые сохраняются в зрелых РНК. 5S-pPHK образуется отдельно.
рРНК как эукариотических, так и прокариотических клеток образуются из более длинных молекул-предшественников, называемых прерибосомными РНК. У прокариот 16S- и 23S-pPHK (гл. 29) образуются из одного длинного 30S- предшественника, молекулярная масса которого составляет приблизительно 2∙106. Этот предшественник метилируется по специфическим основаниям и расщепляется, давая 17S- и 25S-промежуточные РНК, которые затем процессируются путем отщепления остатков с помощью нуклеаз, образуя характерные для прокариот 16S- и 23S-pPHK (рис. 28-19). 5S-pPHK образуется отдельно из 3'-концевого участка 30S-предшественника.
У эукариот 18S- и 28S-pPHK образуются в несколько этапов из большой 45S-прерибосомной РНК. Процессинг 45S-PHK протекает в ядрышке. Сначала происходит метилирование более чем 100 из 14 000 нуклеотидов 45S-предшественника; модификации подвергаются главным образом 2'-гидроксильные группы рибозных остатков. Как показано на рис. 28-20, метилированная 45S-PHK претерпевает затем ряд ферментативных расщеплений, приводящих в конечном итоге к появлению 18S-, 28S- и 5,8S-pPHK, характерных для эукариотических рибосом. 5S-pPHK эукариотов синтезируется отдельно.
тРНК также образуются из более длинных РНК-предшественников в результате ферментативного удаления лишних нуклеотидов с 5'- и 3'-концов молекулы. В некоторых случаях из одной длинной молекулы-предшественника в результате ферментативного расщепления образуются две и даже большее число разных тРНК. Как мы увидим ниже (разд. 29.20), существуют по меньшей мере 32 различные тРНК, а возможно, их гораздо больше.
В ходе посттранскрипционного процессинга в предшественниках тРНК наряду с удалением концевых последовательностей происходят изменения двоякого рода. Во-первых, к некоторым тРНК присоединяется 3'-концевая тринуклеотиднаяпоследовательность —С—С—А (3'); в других тРНК этот 3'-концевой тринуклеотид уже содержится в транскрипте. Ниже мы увидим, что 3'-концевой остаток А представляет собой именно ту часть молекулы тРНК, с которой ковалентно связывается соответствующая ей аминокислота перед включением в растущую полипептидную цепь на рибосоме. Во-вторых, ряд оснований в тРНК специфическим образом модифицируется: одни метилируются, другие дезаминируются, третьи восстанавливаются. Как мы увидим дальше (гл. 29), модифицированные основания располагаются во всех тРНК в определенных положениях.
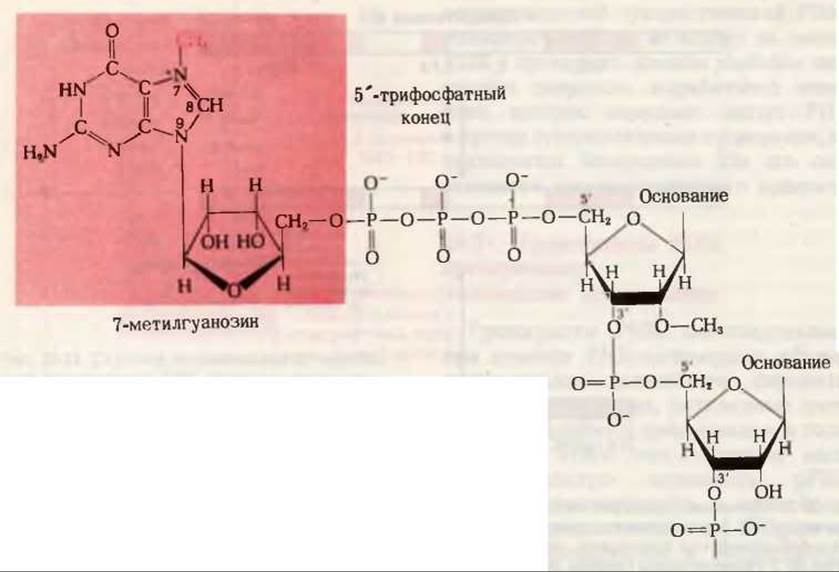
Рис. 28-21. “Кэп”, представляющий собой остаток 7-метилгуанозина и расположенный на 5'-конце эукариотической мРНК; метильная группа выделена красным шрифтом. Обратите внимание на то, что “кэп” соединен с 5'-концевым нуклеотидом трифосфатным мостиком. Почти все эукариотические мРНК содержат 5'- “кэпы”.