Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Синтез белка и его регуляция
Гипотеза оперона
Молекулярные и генетические связи между индукцией и репрессией ферментов прояснились в результате генетических исследований Франсуа Жакоба и Жака Моно из Пастеровского института в Париже. Их классическая работа по индукции ß-галактозидазной активности в клетках Е. coli привела авторов к формулированию гипотезы оперона для объяснения генетического контроля синтеза белка у прокариот. С тех пор эта гипотеза получила полное подтверждение в прямых биохимических экспериментах. Тип регуляции белкового синтеза, рассматриваемый в гипотезе оперона, представляет собой контроль на уровне транскрипции, поскольку регуляция здесь осуществляется главным образом за счет изменения скорости транскрипции генов, т. е. на стадии образования мРНК. Другой основной путь регуляции белкового синтеза - это контроль на уровне трансляции, т. е. регуляция скорости синтеза полипептидной цепи на матрице мРНК. У бактерий ведущая роль в регуляции экспрессии генов принадлежит, по-видимому, контролю на уровне транскрипции. Контроль биосинтеза белка на уровне трансляции, механизм которого остается не совсем понятным, имеет, вероятно, второстепенное значение для бактерий, но он очень важен для эукариот. Кроме этого, в клетках существуют и другие механизмы, которые позволяют выполнять тонкую регулировку скорости синтеза белка.
На основании своих экспериментов Жакоб и Моно предположили, что три структурных гена z, у и а, кодирующие синтез индуцируемых лактозой ферментов ß-галактозидазы, ß-галактозидпермеазы и белка А соответственно, расположены в хромосоме Е. coli рядом (рис. 29-25). Далее они предположили, что в ДНК около этих генов находится другой, ингибиторный участок і, который способен ингибировать транскрипцию трех структурных генов z, у и а. Было постулировано, что участок і представляет собой регуляторный ген (разд. 27.21), кодирующий аминокислотную последовательность регуляторного белка, называвмого репрессором. Когда ген і транскрибируется с образованием соответствующей мРНК, последняя поступает в рибосомы, где на ней, как на матрице, синтезируется репрессор. Белок-репрессор может связываться с другим специфическим участком ДНК, который называется оператором (рис. 29-25). Было высказано предположение, что связывание белка-репрессора с операторным участком ДНК подавляет катализируемый РНК-полимеразой процесс транскрипции трех структурных генов z, у и а, которые кодируют ферменты, индуцируемые ß-галактозидами. В результате из-за отсутствия матрицы синтез этих ферментов подавляется (т. е. репрессируется).
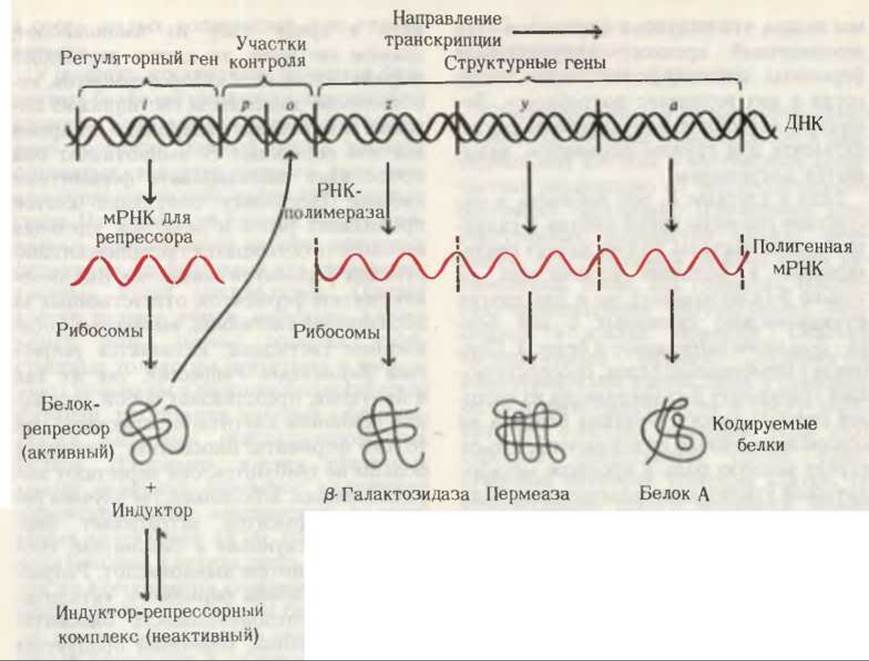
Рис. 29-25. Схематическое изображение lac-оперона. Три структурных lас-гена z, у и а расположены рядом. Перед ними находятся два регуляторных участка р (промотор) и о (оператор). Рисунок дан не в масштабе; участки р и о очень малы по сравнению с генами. Регуляторный ген і кодирует белок-репрессор. Этот белок имеет два центра связывания: один для оператора, другой для индуктора. Активная форма белка-репрессора может присоединяться к оператору, препятствуя тем самым связыванию РНК-полимеразы и последующей транскрипции структурных генов z, у и а. В этих условиях ß-галактозидаза и два других белка клетками не синтезируются. Однако, если в среде вместо глюкозы присутствует лактоза, индуктор соединяется с репрессором, переводя его в неактивное состояние, в котором тот не способен взаимодействовать с оператором. В этом случае РНК-полимераза может связаться с промотором, пройти через зону оператора и начать транскрибировать три структурных гена с образованием полигенной мРНК, которая кодирует синтез трех laс-белков в рибосомах. Более детально функция промотора рассмотрена на рис. 29-27. Лактоза сама по себе не служит индуктором lас-оперона; эту функцию выполняет ее изомер аллолактоза, образующаяся из лактозы.
Чтобы объяснить действие индуктора (случай, когда глюкозы в среде нет, но присутствует лактоза), Жакоб и Моно предположили, что индуктор взаимодействует со вторым специфическим связывающим участком белка-репрессора, т. е. с центром связывания индуктора. При этом образуется индуктор-репрессорный комплекс, что приводит к снижению сродства репрессора к операторному участку ДНК и к освобождению последнего. Как только индуктор-репрессорный комплекс покидает оператор, структурные гены ß-галактозидазы и двух других белков оказываются доступными для транскрипции и РНК-полимераза синтезирует с них мРНК. Эти мРНК используются далее в качестве матриц для синтеза указанных белков в рибосомах, в результате чего клетка получает возможность утилизировать лактозу в качестве источника углерода и энергии.
Предположим теперь, что мы изъяли клетки из среды с лактозой, промыли их и поместили в среду, содержащую вместо лактозы D-глюкозу - субстрат, который клетки всегда способны утилизировать. Поскольку концентрация лактозы в клетке становится при этом исчезающе малой, индуктор, связанный с белком-репрессором, отделяется от него, молекула репрессора возвращается в свое активное состояние и со свойственной ему высокой степенью сродства присоединяется к оператору. Вследствие этого структурные
гены ß-галактозидазы и двух других белков перестают транскрибироваться и из-за отсутствия их мРНК синтез данных белков прекращается. Таким образом, белок-репрессор благодаря своей способности обратимо связываться ю с индуктором, то с оператором (но не одновременно с обоими) может определять как индукцию, так и репрессию синтеза галактозидазы.
Три структурных гена z, у и а вместе с их оператором о были названы Жакобом и Моно опероном, в данном случае lас-опероном (рис. 29-25). Таким образом, в состав оперона входит группа функционально связанных друг с другом структурных генов, которые могут координированно включаться и выключаться, и их оператор. У Е. coli, Salmonella typhimurium и других бактерий было выявлено большое число оперонов (табл. 29-4). Одним из наиболее сложных оперонов является гистидиновый оперон. Он состоит из девяти структурных генов, кодирующих набор ферментов, необходимых для биосинтеза гистидина. Регуляторный ген/lis-оперона кодирует белок-репрессор, который присоединяется к his-оператору и тем самым препятствует транскрипции всех девяти белков оперона в условиях, когда в среде присутствует достаточное количество гистидина.
Таблица 29-4. Некоторые бактериальные опероны
Оперон |
Число ферментных белков |
Функция |
lac |
3 |
Гидролиз и транспорт ß-галактозидов |
his |
9 |
Синтез гистидина |
leu |
4 |
Превращение а-кетоизовалерьяновой кислоты в лейцин |
ara |
4 |
Транспорт и утилизация арабинозы |