Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Еще о генах: репарация, мутации, рекомбинация и клонирование
Гены часто претерпевают рекомбинацию
До настоящего момента мы обсуждали такие изменения генов, которые возникают самопроизвольно, случайно или под действием факторов окружающей среды. Рассмотрим теперь изменения в генах и хромосомах, представляющие собой нормальные события в жизни клеток.
Нормальный биологический обмен между генами или объединение генов из разных источников с образованием измененной хромосомы, способной после этого реплицироваться, транскрибироваться и транслироваться, называется генетической рекомбинацией. Она встречается в различных биологических ситуациях. Мы уже детально познакомились с одним типом генетической рекомбинации - с трансформацией бактерий под действием экзогенной ДНК, которая имела место в классическом эксперименте Эвери, Мак-Леода и Мак-Карти (рис. 27-6). Напомним, что в этом эксперименте ДНК из вирулентного штамма пневмококка попадала в клетки невирулентного штамма и превращала этот штамм в вирулентный. Очевидно, ген вирулентности, присутствующий в ДНК донорной клетки, включается в геном реципиентной клетки. Такая трансформация бактериальных клеток, реализуемая вследствие рекомбинации генов, может наблюдаться не только в лаборатории, но и в естественных условиях.
Другим протекающим в природе процессом генетической рекомбинации является лизогения. При заражении бактериальной клетки определенными видами фагов ДНК этих фагов могут ковалентным образом встраиваться в кольцевую хромосому клетки-хозяина вместо того, чтобы сразу приступить к образованию дочерних фаговых частиц с последующим лизисом клеток, как это бывает в случае обычной фаговой инфекции. Встроившись в хромосому клетки-хозяина, фаговый геном может реплицироваться в ее составе в течение многих поколений, не проявляя себя в форме новых фаговых частиц. Однако спустя некоторое время какое-нибудь событие может “включить” механизм экспрессии этих “спящих” фаговых генов, в результате чего начнется образование фаговых частиц и наступит лизис клетки-хозяина (рис. 30-11). Фаги, ДНК которых может встраиваться в хромосому клетки-хозяина и существовать там в неэкпрессируемом виде, называются умеренными, или лизогенизирующими фагами. Наиболее известный умеренный фаг - это фаг λ; его включение в хромосому Е. coli изучено очень подробно. Считается, что такой же тип генетической рекомбинации имеет место при заражении человека вирусом простого герпеса, который вызывает герпетическую лихорадку, а также воспаление и язвы половых органов. ДНК вируса простого герпеса способна встраиваться в геном клеток человека и находиться там в бездействии до тех пор, пока какое-нибудь событие не “включит” механизм образования инфекционных вирусных частиц.
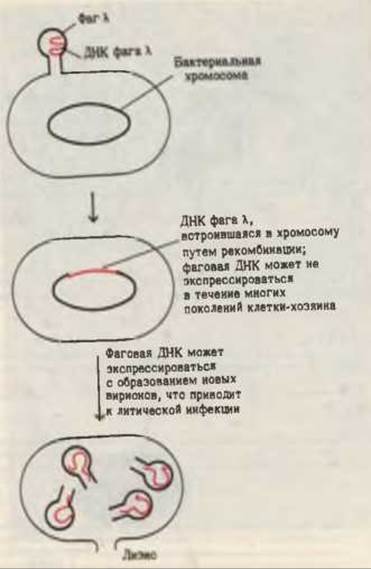
Рис. 30-11. Встраивание ДНК фага λ в хромосому Е. coli в неэкспрессируемом состоянии, которое может поддерживаться посредством репликации в течение многих поколений. В результате некоего события, играющего роль пускового механизма, вирусный геном может начать экспрессироваться с образованием фаговых частиц и последующим лизисом клеток.
Еще одним типом генетической рекомбинации является трансдукция (рис. 30-12). Если бактериальная клетка заражена некоторыми ДНК-содержащими фагами, то небольшая часть ее хромосомы может ковалентно присоединиться к фаговой ДНК, реплицироваться вместе с ней и таким образом встраиваться в ДНК дочерних фаговых частиц. Когда такие частицы заражают другую клетку, фаговая ДНК приносит в эту клетку участок хромосомы первой клетки. Трансдукция (что означает “перенос”) - это природный процесс, который в лабораторных условиях используется для картирования бактериальных хромосом.
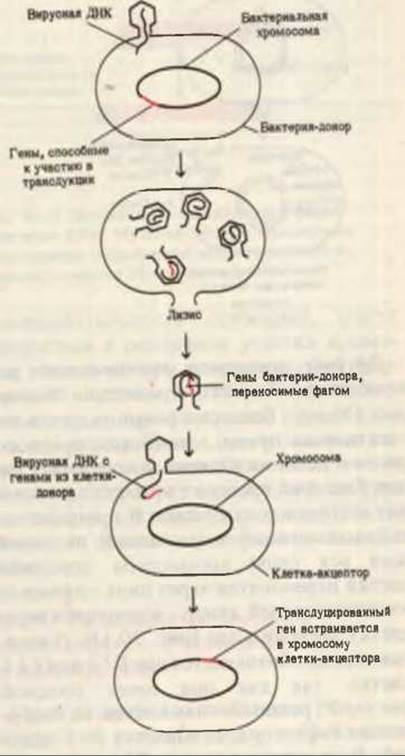
Рис. 30-12. Генетическая рекомбинация в ходе вирусной трансдукции бактериальных генов в реципиентную клетку.
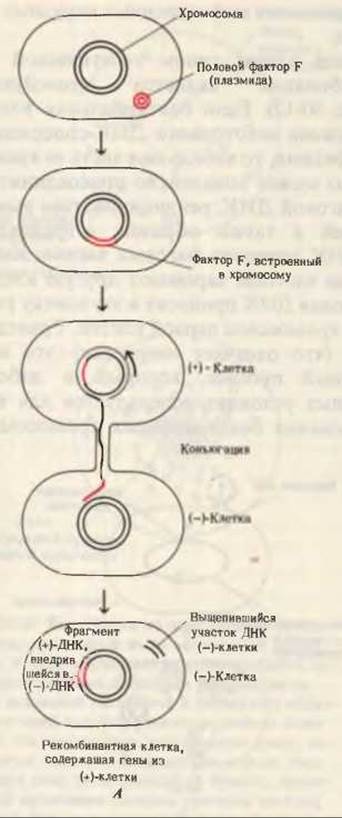
Другим примером генетической рекомбинации служит конъюгация бактерий. Обычно бактерии размножаются вегетативным путем, с помощью простого роста и деления. Однако у некоторых видов бактерий время от времени происходит половая конъюгация. В процессе такой конъюгации часть одной из цепей (или вся цепь) хромосомы донорной клетки переносится через пиль - длинный соединительный канал - в реципиентную клетку того же вида (рис. 30-13). Донорная клетка обозначается как F+-, или (+)-клетка, так как она несет половой фактор F; реципиентная клетка, не содержащая F-фактора, называется (-) - клеткой. В результате половой конъюгации реципиентная клетка приобретает несколько новых генов, которые встраиваются в ее хромосому.

Рис. 30-13. А. Перенос и рекомбинация генов в процессе конъюгации бактерий. ДНК (+)-клетки реплицируется по механизму “катящегося кольца”, и образующаяся одиночная цепь, содержащая фактор F, вводится в (—)-клетку. Б. Электронная микрофотография конъюгирующих клеток Е. coli. (+)-Клетка, расположенная в верхней части микрофотографии, соединена с (—)-клеткой с помощью одного длинного пиля.
У эукариотических организмов генетическая рекомбинация осуществляется при половом слиянии яйцеклетки и сперматозоида: появляющиеся в потомстве клетки содержат дочерние хромосомы, состоящие из определенных генов обеих родительских хромосом (рис. 30-14). В этом процессе хромосомы сперматозоида и яйцеклетки расщепляются в гомологичных точках, а затем куски хромосом двух родительских клеток обмениваются своими генами и соединяются с образованием новых комбинаций генов. В результате потомство слившихся клеток обладает комбинацией фенотипических признаков, принадлежавших обоим родителям. Этот природный процесс расщепления, сборки и соединения генов и наборов генов в ходе половой конъюгации эукариот протекает с высокой точностью, без нарушения рамки считывания и сигнальных последовательностей в ДНК.
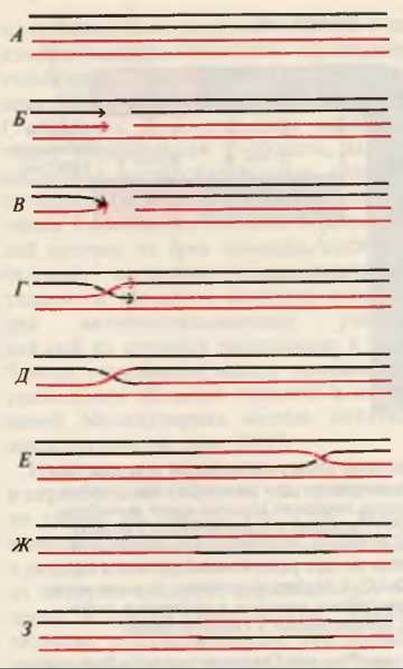
Рис. 30-14. Схематическое изображение рекомбинации генов между двумя гомологичными родительскими ДНК [они показаны на рис. (Л) красным и черным цветом], согласно модели “одноцепочечного переключения”. Одна цепь каждой из ДНК расщепляется (Б), “переключается” на другую ДНК (В, Г) и присоединяется к противоположной расщепленной цепи (Д). Обмен цепями между молекулами ДНК распространяется вдоль хромосомы (Е) до определенной точки, в которой вновь происходит расщепление цепей (Ж) и их обратное “переключение” (3), т. е. рекомбинация завершается.