Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Еще о генах: репарация, мутации, рекомбинация и клонирование
Выделение генов и получение кДНК
Из фрагментов вирусных и бактериальных хромосом уже выделен целый ряд генов. Что же касается выделения специфических генов из фрагментированных эукариотических хромосом, то реализация этой процедуры все еще остается довольно сложной и трудоемкой задачей. Существует два основных подхода для получения специфических генов, подлежащих затем рекомбинации и клонированию. В одном из них, который получил название “шотган” (от англ. shotgun - дробовик), всю клеточную ДНК обрабатывают рестриктирующей эндонуклеазой, образующей в местах разрыва выступающие концы. Полученные фрагменты ДНК встраивают затем в плазмиды Е. coli, “раскрытые” (т. е. переведенные в линейную форму) с помощью той же самой рестриктирующей эндонуклеазы. В результате образуется чрезвычайно сложная смесь, состоящая, вероятно, из тысяч разных рекомбинантных плазмид, среди которых лишь одна может содержать нужный ген. Для поиска плазмиды, несущей этот ген, разработаны специальные процедуры, которые называют скринингом. Одна из таких процедур описана в разд. 30.17.
Другой подход, используемый для получения нужных генов, состоит в конструировании на мРНК-матрице комплементарной по отношению к ней ДНК (кДНК). Хотя, как мы уже говорили, большинство клеток содержит трудноразделимые смеси множества различных мРНК, тем не менее иногда удается выделить чистую мРНК, кодирующую какой-либо один специфический белок. Для этого используют специализированные клетки, которые вырабатывают преимущественно какой-то один вид белка. Например, из ретикулоцитов - незрелых красных кровяных клеток-, в которых гемоглобин составляет 90% синтезируемого белка, можно выделить мРНК для а- и ß-полипептидных цепей гемоглобина. Аналогичным образом из В-клеток островков Лангерганса поджелудочной железы можно выделить мРНК для проинсулина человека (разд. 25.11).
Однако более общий подход к получению мРНК, кодирующей специфический белок, состоит в следующем. Клетки лизируют, собирают центрифугированием полисомы и обрабатывают их антителами против того белка, ген которого хотят извлечь. В популяции полисом должны присутствовать полисомы, синтезирующие данный белок на его матрице мРНК. В таких полисомах интересующий нас белок находится на разных стадиях своего синтеза (разд. 29.14). Очевидно, специфические антитела будут взаимодействовать только с полностью (или почти полностью) достроенным белком, который еще прикреплен к полисомам, синтезирующим его на матрице мРНК. Специфический комплекс антител с полисомами будет при этом выпадать в осадок и таким образом отделяться от прочих полисом. Специфическую мРНК, на которой синтезировался белок, можно затем извлечь из осадка и выделить с помощью хроматографических методов в практически чистом виде, т. е. без примесей других мРНК.
Теперь специфическую мРНК, кодирующую белок, ген которого надо получить, можно использовать в качестве матрицы для ферментативного синтеза кДНК с помощью обратной транскриптазы (разд. 28.26). Однако для функционирования обратной транскриптазы необходима затравочная ДНК. Напомним (разд. 28.22), что на 3'-конце молекул мРНК находится poly (А)-последовательность. Поэтому к мРНК добавляют poly(T), которая образует с роlу(А)-хвостом мРНК двухцепочечные участки (рис. 30-21), служащие затравкой для обратной транскриптазы. В таких условиях обратная транскриптаза синтезирует на матрице мРНК комплементарную цепь кДНК из смеси dATP, dTTP, dGTP и dCTP. Затем мРНК удаляют из гибрида мРНК-кДНК и на одноцепочечной кДНК с помощью ДНК-полимеразы I достраивают вторую цепь; в результате образуется двухцепочечная кДНК, содержащая “шпильку”. После расщепления “шпильки” (рис. 30-21) остается синтетическая двухцепочечная кДНК, соответствующая белку, кодируемому интересующим нас геном. Чтобы упростить последующие процедуры, кДНК в ходе реакции с обратной транскриптазой обычно синтезируют из 32Р-дезоксирибонуклеозид-5'-трифосфатов.
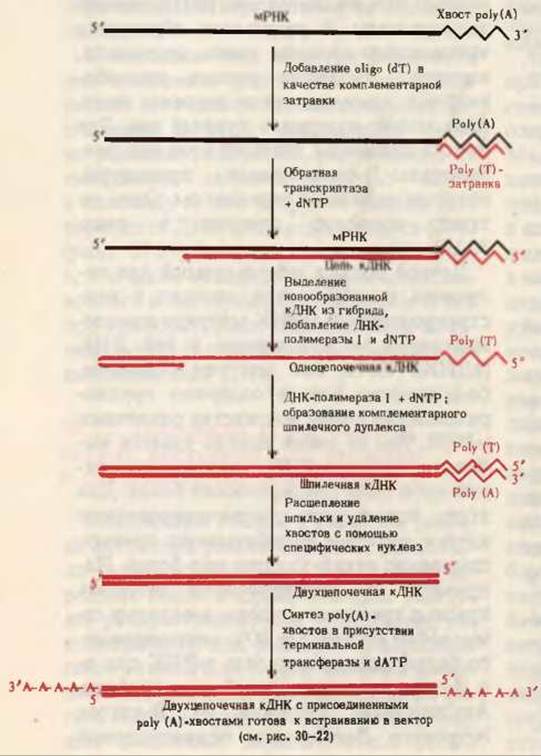
Рис. 30-21. Конструирование двухцепочечной кДНК на основе мРНК. Выделенную мРНК данного гена транскрибируют с помощью обратной транскриптазы с образованием кДНК. Для работы обратной транскриптазы необходима затравочная ДНК, роль которой выполняет poly(T), комплементарная 3'-концевому роlу(А)-хвосту мРНК. Полученную цепь кДНК, которую обычно метят с помощью радиоактивных предшественников dNTP, отделяют и используют одновременно в качестве матрицы и в качестве затравки для синтеза двухцепочечной шпилечной кДНК в присутствии ДНК-полимеразы I. Затем “шпильку” расщепляют, цепи подравнивают и к полученной двухцепочечной кДНК с тупыми концами присоединяют роlу(А)-хвосты (рис. 30-20). Теперь такая кДНК, несущая poly (А), может быть встроена в вектор (рис. 30-22). Для обнаружения соответствующего природного гена во фрагментированных хромосомах применяют радиоактивную кДНК.
В результате мы имеем синтетическую кДНК, которая кодирует аминокислотную последовательность данного белка. Заметим, однако, что если эту синтетическую кДНК получали с эукариотической мРНК, то она не идентична природному гену этого белка, поскольку не содержит ни нитронов, т. е. вставочных последовательностей, ни стартовых и терминирующих сигналов, присущих генам большинства эукариотических белков.