Основы биохимии Том 3 - А. Ленинджер 1985
Некоторые аспекты биохимии человека
Пищеварение, транспорт питательных веществ и взаимосвязь обменных процессов
Скелетные мышцы используют АТР для выполнения по мере надобности механической работы
На долю скелетных мышц в целом приходится более 50% кислорода, потребляемого организмом человека в состоянии покоя, и до 90%-при интенсивной мышечной работе. Обмен веществ в скелетных мышцах направлен в первую очередь на выработку АТР как непосредственного источника энергии для сокращения и расслабления. Кроме того, скелетные мышцы приспособлены к тому, чтобы выполнять механическую работу не непрерывно, а по мере надобности. Иногда, например при беге спринтера на 100 м, скелетные мышцы выполняют огромную работу за очень короткое время.
В качестве топлива скелетные мышцы в зависимости от степени их активности используют глюкозу, свободные жирные кислоты или кетоновые тела. В покоящихся мышцах основными субстратами энергетического обмена служат свободные жирные кислоты и кетоновые тела, доставляемые с кровью из печени. Эти субстраты подвергаются окислению и распаду до ацетил-СоА, который вступает далее в цикл лимонной кислоты и окисляется до СO2. Сопутствующий перенос электронов к кислороду обеспечивает энергией процесс окислительного фосфорилирования и превращение ADP в АТР. При умеренной нагрузке в дополнение к жирным кислотам и кетоновым телам мышцы используют еще и глюкозу крови. При этом глюкоза подвергается фосфорилированию и распадается в ходе гликолиза до пирувата, который далее через ацетил-СоА окисляется в цикле лимонной кислоты. Наконец, при максимальной мышечной нагрузке расход АТР на сокращение настолько велик, что скорость доставки субстратов (топлива) и кислорода кровью оказывается недостаточной. В этих условиях в ход идет накопленный в самих мышцах гликоген, который расщепляется до лактата путем анаэробного гликолиза; при этом на один расщепившийся остаток глюкозы образуются две молекулы АТР (разд. 15.7,ж). Таким образом, анаэробный гликолиз дает дополнительное количество АТР сверх того основного количества, которое вырабатывается при аэробном окислении других субстратов энергетического обмена в цикле лимонной кислоты. Использование глюкозы крови и гликогена мышц как срочно мобилизуемого топлива для мышечной работы резко возрастает при увеличении секреции адреналина, который в печени стимулирует образование из гликогена глюкозы, поступающей в кровь, а в мышечной ткани расщепление гликогена до лактата (гл. 25). Поскольку в скелетных мышцах нет глюкозо-6-фосфатазы, мышечный гликоген предназначен только для выработки энергии гликолитическим путем.
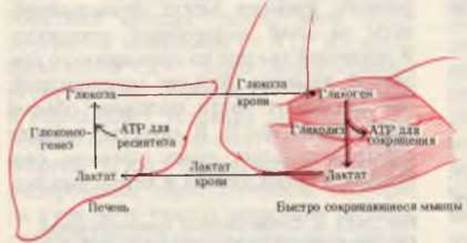
Рис. 24-12. Взаимосвязь обмена веществ в скелетных мышцах и в печени. При тяжелой физической нагрузке источником энергии для скелетных мышц служит гликоген, распадающийся гликолитическим путем. В восстановительный период часть образовавшегося в мышцах лактата переносится в печень и превращается в глюкозу, которая поступает в кровь и доставляется в мышцы, где используется для восполнения запасов гликогена.
Запасы гликогена в мышцах, однако, невелики, и потому существует верхний предел того количества энергии, которое вырабатывается в ходе гликолиза в условиях максимальной (например, при спринте) нагрузки. Более того, накопление молочной кислоты и связанное с этим снижение pH, а также повышение температуры, происходящее при очень высокой мышечной активности, снижают эффективность обмена в мышцах. Так, в период восстановления после максимальной мышечной нагрузки атлет продолжает еще некоторое время тяжело дышать. Потребляемый при этом дополнительный кислород используется для окисления пирувата, лактата и других субстратов, а также регенерации АТР и фосфокреатина в мышцах. Одновременно лактат крови превращается в печени путем глюконеогенеза в поступающую в кровь глюкозу, которая попадает в мышцы и используется на восстановление запасов гликогена (разд. 20.1). Таким образом, дополнительно потребленный кислород (“кислородный долг”) восстанавливает нормальное метаболическое состояние организма посредством целой серии процессов, в ходе которых происходит взаимодействие мышц и печени (рис. 24-12).
Есть и другой путь, который обеспечивает скелетные мышцы максимальным количеством АТР в критических обстоятельствах. В мышцах содержится значительное количество фосфокреатина (разд. 14.15), который с помощью креатинкиназы способен быстро восполнять отцепившиеся при мышечном сокращении концевые фосфатные группы АТР:
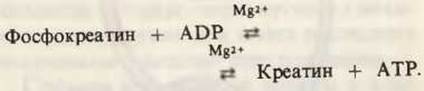
В периоды высокой сократительной активности и интенсивного гликолиза реакция сильно смещена вправо, тогда как в период восстановления происходит ресинтез фосфокреатина из креатина за счет АТР.
АТР в скелетных мышцах нужен не только для того, чтобы обеспечивать скольжение нитей актина вдоль нитей миозина, или толстых нитей (разд. 14.14), но также и для расслабления. Мышечное сокращение инициируется импульсом, поступающим от двигательного нерва; этот импульс передается на поперечные трубочки и на саркоплазматический ретикулум, из которого в саркоплазму выходят ионы Са2+. Далее Са2+ связывается с тропонином - регуляторным белком, который преобразует этот сигнал
в процесс скольжения актиновых нитей за счет энергии АТР. После прекращения подачи импульсов со стороны двигательного нерва ионы Са2+ должны быть удалены из саркоплазмы, чтобы могло произойти расслабление мышцы. Это достигается транспортом ионов Са2+ обратно в саркоплазматический ретикулум с помощью Са2+ -транспортирующей мембранной АТРазы. Перенос двух ионов Са2+ внутрь саркоплазматического ретикулума происходит за счет гидролиза одной молекулы АТР, т. е. на расслабление скелетной мышцы тратится почти столько же энергии, сколько на ее сокращение.