Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
ДНК: структура хромосом и генов
Нуклеотидная последовательность ДНК выполняет функцию матрицы
Молекулы ДНК представляют собой длинные полимерные цепи, имеющие специфические последовательности четырех главных оснований А, Т, G и С, которые являются символами, предназначенными для кодирования генетической информации. Поэтому мы говорим, что нуклеотидная последовательность в ДНК служит матрицей при репликации ДНК. Однако важно понять, почему для точной репликации, транскрипции и трансляции генетической информации нам необходимы матрицы.
При биосинтезе неинформационной макромолекулы гликогена, которая состоит из повторяющихся единиц только одного типа-D-глюкозы, тождественность и чистота конечного продукта обеспечиваются активным центром гликоген-синтазы (разд. 20.13). Для этого фермента характерна субстратная специфичность, т. е. его активный центр способен присоединять только молекулу UDP-глюкозы и нередуцирующий конец цепи молекулы гликогена, которая должна быть удлинена. В принципе активный центр этого фермента (как, впрочем, и всех других ферментов) можно рассматривать как матрицу (это слово означает “шаблон” или “форму”), поскольку между молекулой (или молекулами) субстрата и активным центром осуществляется комплементарная “подгонка”.
В случае ДНК, РНК и полипептидов только один активный центр фермента не в состоянии обеспечить специфическую последовательность кодирующих единиц. Активные центры ферментов сравнительно малы и могут связывать одновременно лишь одну или несколько молекул, играющих роль строительных блоков, в таком положении, чтобы обеспечить их точную сборку в правильной последовательности. Нуклеиновые же кислоты, содержащие в своем составе тысячи или миллионы нуклеотидных единиц, настолько велики, что размеры активных центров ферментов оказываются просто недостаточными для того, чтобы точно определять полную последовательность, в которой должны быть собраны нуклеотидные единицы. Следовательно, матрицей для формирования нуклеотидной последовательности своего комплементарного партнера должна служить одна цепь ДНК.
Другой аспект гипотезы Уотсона Крика состоит в том, что структура двойной спирали ДНК указывает способ, с помощью которого может быть точно воспроизведена содержащаяся в ДНК генетическая информация (рис. 27-13). Поскольку две цепи двойной спирали ДНК структурно комплементарны, их нуклеотидные последовательности несут комплементарную друг по отношению к другу информацию. Уотсон и Крик постулировали, что репликация ДНК в ходе деления клеток начинается с разделения двух цепей, каждая из которых становится матрицей, определяющей нуклеотидную последовательность новой комплементарной цепи, образуемой с помощью репликативных ферментов. Была высказана мысль, что правильность репликации каждой из цепей ДНК должна обеспечиваться точным соответствием и стабильностью комплементарных пар оснований А=Т и G=C в двух дочерних дуплексах, каждый из которых содержит одну цепь родительской ДНК и новую цепь, комплементарную этой родительской цепи. Было постулировано также, что каждая вновь образованная дочерняя двойная спираль попадает в дочернюю клетку без каких-либо изменений. В гл. 28 мы увидим, как эта гипотеза была экспериментально подтверждена.
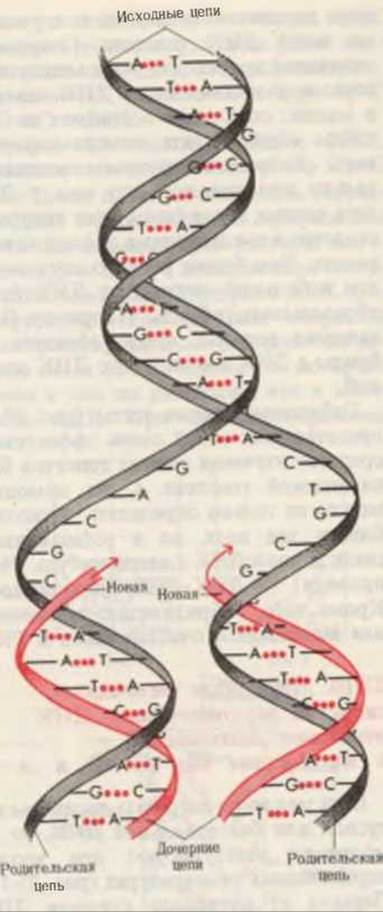
Рис. 27-13. Модель репликации ДНК, предложенная Уотсоном и Криком. Комплементарные цепи родительской ДНК разделяются, и каждая из них служит матрицей для биосинтеза комплементарной дочерней цепи (дочерние цепи показаны красным цветом).