Биохимия человека Том 2 - Марри Р. 1993
Биохимия внутри- и межклеточных коммуникаций
Гормоны половых желез
Регуляция и физиологическое действие гормонов яичников
Созревание и поддержание функции женской репродуктивной системы
Основная функция яичниковых гормонов — подготовка структурных компонентов женской половой системы (см. ниже) к размножению. Эта подготовка включает 1) созревание примордиальных зародышевых клеток; 2) развитие тканей, необходимых для имплантации бластоцисты; 3) обеспечение гормонального контроля времени овуляции; 4) установление с помощью плацентарных гормонов среды, необходимой для поддержания беременности и 5) обеспечение гормональной регуляции родов и лактации.
Эстрогены стимулируют развитие тканей, участвующих в размножении. Как правило, под влиянием этих гормонов повышается скорость синтеза белка, рРНК, тРНК, мРНК и ДНК, что приводит к увеличению размеров и числа клеток соответствующих тканей. Эстрогенная стимуляция обусловливает пролиферацию и дифференцировку влагалищного эпителия, пролиферацию эндометрия, а также гипертрофию с увеличением длины его желез; появление собственной ритмической подвижности миометрия; пролиферацию протоков грудных желез. Эстрадиол оказывает также анаболическое действие на кости и хрящи, способствуя таким образом росту. Воздействуя на периферические кровеносные сосуды, эстрогены обычно вызывают их расширение и усиливают теплоотдачу.
Для проявления активности прогестинов обычно требуется предшествующее или одновременное действие эстрогенов. Таким образом, гормоны двух этих классов часто функционируют синергично, хотя могут быть и антагонистами. Прогестины уменьшают стимулирующее действие эстрогенов на пролиферацию эпителия влагалища и способствуют переходу эпителия матки из пролиферативной фазы в секреторную (увеличение размеров и функции секреторных желез и повышение содержания гликогена), подготавливая его к имплантации оплодотворенной яйцеклетки. Эти гормоны усиливают развитие ацинарной части грудных желез после эстрогенной стимуляции развития протоков. Прогестины снижают периферический кровоток, уменьшая тем самым теплопотерю. В результате в лютеиновой фазе менструального цикла, на которую приходится образование данного гормона, температура тела повышается. Такие температурные скачки, составляющие обычно около 0,5°С, используются в качестве показателя овуляции.
Максимальное число оогониев в яичниках плода человека достигает 6—7 млн. примерно к пятому месяцу эмбрионального развития. К моменту рождения оно понижается примерно до 2 млн., а после наступления менархе составляет 100000—200000. Примерно 400—500 из них развиваются в зрелые ооциты. Остальные постепенно исчезают в результате какого-то пока еще не раскрытого процесса. Известно лишь, что в этом процессе участвуют яичниковые андрогены. Созревание фолликулов начинается в младенческие годы; на протяжении всего препубертатного периода яичники увеличиваются в размерах в результате увеличения объема фолликулов, обусловленного ростом клеток гранулезы, накопления ткани атрезированных фолликулов и увеличения массы медуллярной стромальной ткани с клетками интестициальной ткани и теки, способными продуцировать стероидные гормоны.
В детстве концентрация половых гормонов низка, хотя экзогенные гонадотропины увеличивают их продукцию. Следовательно, незрелые яичники обладают способностью синтезировать эстроген. Существует предположение, что у неполовозрелых девочек имеющиеся у них в небольшом количестве половые стероиды подавляют образование гонадотропинов, а в пубертатном возрасте гипоталамогипофизарная система становится менее чувствительной к ингибирующему действию этих гормонов. В период полового созревания начинается импульсная секреция ГнРГ (гонадолиберина); под влиянием ЛГ резко повышается уровень образования яичниковых гормонов, а под влиянием ФСГ, главного стимулятора секреции эстрогенов, происходит созревание фолликулов и наступает овуляция.
Менструальный цикл
Частота овуляции и готовность к половому акту определяются гормонами. У видов с моноэстральным циклом овуляция и спаривание происходят раз в году, у видов с полиэстральным циклом овуляция повторяется несколько раз в году. У приматов, для которых характерны менструальные циклы с отторжением эндометрия в конце каждого цикла, половое поведение не имеет тесной связи с овуляцией. У человека менструальный цикл обусловливается сложным взаимодействием между гипоталамусом, гипофизом и яичниками. В норме продолжительность менструального цикла варьирует от 25 до 35 дней (в среднем 28 дней). Его можно подразделить на фолликулярную фазу, лютеиновую фазу и менструацию (рис. 50.8).
А. Фолликулярная фаза. По каким-то невыясненным причинам под влиянием ФСГ начинает увеличиваться лишь один из фолликулов. В первую неделю фолликулярной фазы содержание Е2 остается низким, но затем по мере увеличения фолликула начинает прогрессивно повышаться. За 24 ч до ника ЛГ (ФСГ) уровень Е2 достигает максимума и сенсибилизирует гипофиз к действию гонадолиберина (ГнРГ).

Рис. 50.8. Гормональные и физиологические изменения во время типичного менструального цикла у человека. М — менструация, ЛГ — лютеинизирующий гормон, ФСГ — фолликулостимулирующий гормон. (Reproduced, with permission, from Midgley A. R. In: Human Reproduction, Hafez ESE, Evans TN [editors]. Harper and Row, 1973.)
Выброс ЛГ обусловливается либо этим высоким уровнем Е2 по механизму «положительной обратной связи», либо резким падением его уровня. Продолжительное введение высоких доз эстрогенов (в качестве пероральных контрацептивов) снижает как секрецию ЛГ и ФСГ, так и действие ГнРГ на гипофиз. Содержание прогестерона в фолликулярной фазе очень мало.
Б. Лютеиновая фаза. После овуляции клетки гранулезы лопнувшего фолликула лютеинизируются и образуют желтое тело — структуру, которая вскоре начинает вырабатывать прогестерон и некоторое количество эстрадиола. Уровень эстрадиола достигает максимума примерно к середине лютеиновой фазы, а затем резко снижается. Основной гормон лютеиновой фазы цикла — прогестерон. Он необходим для формирования секреторного эндометрия, обеспечивающего нужные условия для развития имплантировавшейся бластоцисты. На первых этапах сохранение желтого тела требует присутствия ЛГ, и гипофиз в течение примерно десяти дней выделяет этот гормон. Если имплантация произошла (22—24-й день цикла), функцию ЛГ берет на себя хорионический гонадотропин (ХГЧ) — плацентарный гормон, очень близкий к ЛГ, вырабатываемый цитотрофобластными клетками имплантированного эмбриона на ранних стадиях развития (см. гл. 45). ХГЧ поддерживает синтез прогестерона желтым телом до тех пор, пока плацента не начнет продуцировать большие количества этого стероида. В отсутствие имплантации (и ХГЧ) желтое тело деградирует и наступает менструация. После отторжения эндометрия начинается новый цикл. Лютеиновая фаза всегда длится 14 + 2 дней. Колебания продолжительности цикла почти во всех случаях обусловлены различиями в фолликулярной фазе.
Беременность и плацентарные гормоны
Имплантированная бластоциста образует трофобласт, который впоследствии организуется в плаценту. Именно плацента обеспечивает связь между системами кровообращения зародыша и матери и вырабатывает ряд гормонов.
А. Хорионический гонадотропин человека (ХГЧ).
Главная функция этого гликопротеинового гормона состоит в том, чтобы поддерживать существование желтого тела до тех пор, пока плацента не начнет продуцировать прогестерон в количествах, достаточных для нормального течения беременности. ХГЧ обнаруживается уже через несколько дней после имплантации, что используется для ранней диагностики беременности. Содержание этого гормона достигает максимума к середине первого триместра, постепенно снижаясь на протяжении остальной беременности. Уровень ХГЧ и других гормонов при беременности показан на рис. 50.9.
Б. Прогестины. В первые 6—8 нед беременности главным источником прогестерона служит желтое тело, затем эту функцию берет на себя плацента. Желтое тело при этом продолжает функционировать, однако на поздних стадиях беременности плацента вырабатывает в 30-40 раз больше прогестерона, чем желтое тело. Поскольку плацента не способна синтезировать холестерол, она должна получать его из материнского организма.

Рис. 50.9. Содержание гормонов во время нормальной беременности. ХГЧ — хорионический гонадотропин человека, ХСЧ — хорионический соматомаммотропин человека (использованы данные различных авторов). (Reproduced, with permission, from Ganong W. F. Review of Medical Physiology, 13th ed. Appleton and Lange, 1987.)
В. Эстрогены. При беременности концентрация эстрадиола, эстрона и эстриола в плазме постепенно повышается. В наибольшем количестве образуется эстриол; его образование отражает ряд фетоплацентарных функций. Надпочечники плода продуцируют дегидроэпиандростерон (ДГЭА) и ДГЭА-сульфат, превращающиеся в печени плода в 16а-гидроксипроизводные, которые в свою очередь преобразуются в плаценте в эстриол. Образующийся эстриол поступает с кровью в печень матери, где конъюгирует с глюкуронидами и в таком виде выделяется с мочой (рис. 50.10). Содержание в моче эстриола используется в качестве показателя для оценки течения ряда процессов у плода и матери.
Не менее интересен обмен субстратами, необходимыми для образования у плода кортизола и ДГЭА. Из-за отсутствия в эмбриональных надпочечниках комплекса 3ß-гидроксистероид-дегидрогеназы/∆5,4-изомеразы синтез кортизола у плода осуществляется за счет плацентарного прогестерона. Прегненолон, необходимый для синтеза дегидроэпиандростерона, также поступает из плаценты (рис. 50.10).
Г. Плацентарные лактогены. Плацента вырабатывает плацентарный лак гоген (ПЛ) — гормон, называемый также хорионическим соматомаммотропином или плацентарным гормоном роста, так как он обладает биологическими свойствами пролактина и гормона роста. Генетическая взаимосвязь этих гормонов рассматривается в гл. 45. Физиологическая роль плацентарного лактогена точно не установлена, поскольку у женщин, лишенных этого гормона, беременность протекает нормально и рождаются здоровые дети.
Роды
Беременность продолжается строго определенное число дней, специфичное для каждого вида. Однако фактор, отвечающий за окончание беременности, не установлен. Предполагают, что важную роль в этом играют гормоны, возможно эстрогены и прогестины, поскольку они влияют на сокращение матки. Имеются данные и об участии катехоламинов в индукции родов. Окситоцин стимулирует сокращение матки, и в клинике его используют для облегчения родов. Однако этот гормон не инициирует роды до окончания беременности. Содержание окситоциновых рецепторов в матке к концу беременности резко увеличивается, превышая в 100 раз их содержание в начале беременности. Это увеличение коррелирует с увеличением к концу беременности количества эстрогенов, под влиянием которых повышается количество окситоциновых рецепторов (гл. 45). При наступлении родов шейка матки растягивается, вызывая рефлекторную стимуляцию высвобождения окситоцина, а следовательно, и дальнейшее сокращение матки. Важную роль в этом процессе могут играть механические факторы, такие, как растяжение или давление на мышцу. При родах у матери и новорожденного резко изменяется гормональная среда, и после отторжения плаценты содержание в плазме прогестерона (определяемого в виде прегнандиола) и эстриола быстро уменьшается (рис. 50.9).

Рис. 50.10. Метаболизм стероидов в системе мать — плод. ДГЭА — дегидроэпиандростерон.
Развитие молочной железы и лактация
Дифференцировка и функция молочной железы регулируются согласованным действием нескольких гормонов. Инициируют этот процесс женские половые гормоны: эстрогены отвечают за рост протоков, а прогестины стимулируют пролиферацию альвеол. Некоторое разрастание железистой ткани, сопровождающееся отложением жировой ткани, происходит при половом созревании, однако наибольшего развития железа достигает при беременности, когда железистая ткань подвергается воздействию высоких концентраций эстрадиола и прогестерона. Как показали исследования, проведенные главным образом на эксплантатах молочных желез крысы, для полной дифференцировки требуется также действие пролактина, глюкокортикоидов, инсулина или фактора роста и какого-то неидентифицированного фактора сыворотки. Из этих гормонов только концентрация пролактина претерпевает при беременности резкие изменения: на поздних сроках беременности она увеличивается от менее 2 нг% до более 200 нг%. Подробно изучено влияние гормонов на синтез различных белков молока, включая лактальбумин, лактоглобулин и казеин. Эти гормоны повышают скорость синтеза перечисленных белков путем увеличения количества специфических мРНК, что, по крайней мере в случае казеина, обусловлено усилением транскрипции гена. Заслуживает внимания тот факт, что усиление транскрипции гена наблюдалось лишь в том случае, если к культуре эксплантата добавляли одновременно кортизол, пролактин и инсулин.
На поздних стадиях беременности образование и секреция молока подавляются под действием прогестерона, необходимого для дифференцировки альвеол. Лактация начинается только после окончания родов, когда содержание этого гормона резко уменьшается. Содержание пролактина после родов также быстро уменьшается, но возрастает при каждом акте кормления (гл. 45), поддерживая таким образом непрерывную лактацию. Если ребенка по каким-либо причинам перестают прикладывать к груди, лактация постепенно прекращается. Парентеральным введением больших доз андрогенов до начала вскармливания можно вызвать быстрое прекращение лактации.
Кормление грудью индуцирует также секрецию окситоцина задней долей гипофиза. Окситоцин стимулирует сокращение миоэпителиальных клеток, окружающих альвеолярные протоки, способствуя таким образом выбросу молока из железы. Механизм регуляции синтеза и секреции окситоцина рассматривается в гл. 45.
Менопауза
У женщин западного полушария в возрасте примерно 53 лет менструальные циклы становятся все более нерегулярными; одновременно из яичников исчезают фолликулы и прекращается их функция. Других источников прогестерона в организме нет, но в результате ароматизации надпочечникового стероида андростендиона образуются значительные количества эстрона, обладающего слабой эстрогенной активностью (рис. 50.6). Количество образующегося эстрона не столь велико, чтобы снизить содержание гипофизарных гонадотропинов, и для постменопаузального периода характерно резкое повышение уровней ЛГ и ФСГ. У женщин в постменопаузе особенно часто возникают две проблемы, связанные с тканевым метаболизмом. Первая из них состоит в том, что эстрон не всегда способен предотвратить атрофию вторичных половых тканей, особенно эпителия нижнего отдела мочеполового тракта и влагалища. Вторая проблема, с которой сталкиваются пожилые женщины, — это остеопороз. У женщин с особенно сильным уменьшением костной массы содержание эстрона оказывается ниже нормы.
Синтетические агонисты и антагонисты
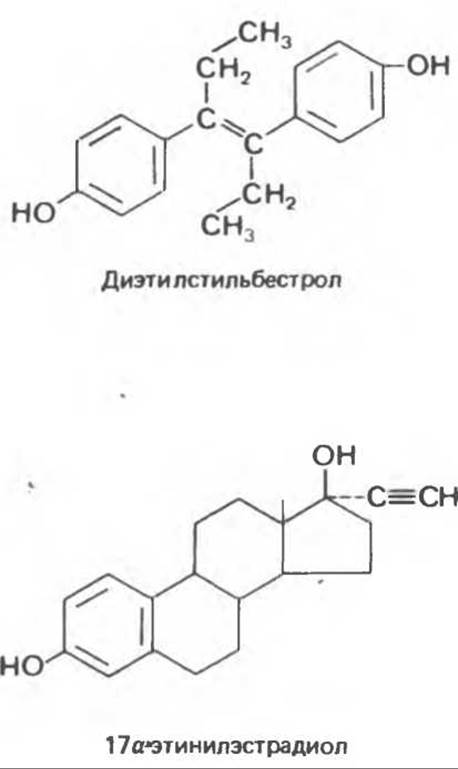
А. Эстрогены. Некоторые синтетические соединения обладают эстрогенной активностью и при этом имеют фармакологические преимущества. Большинство модификаций направлено на ослабление метаболизма в печени, чтобы препараты можно было принимать перорально. Одним из первых синтетических эстрогенов был диэтилстильбестрол. В качестве примеров модифицированных стероидов можно привести также 17а-этинилэстрадиол и местранол, используемые в качестве пероральных контрацептивов.

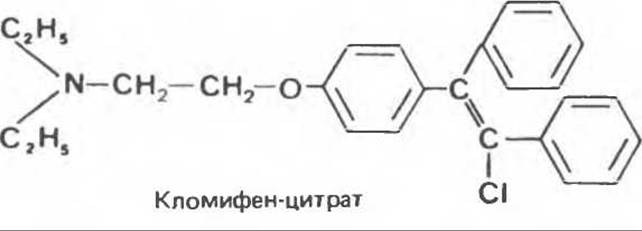
Синтезированы также многочисленные соединения с антиэстрогенной активностью. Некоторые из них нашли клиническое применение. Большинство таких антагонистов действуют, конкурируя с эстрадиолом за его внутриклеточные рецепторы (см. ниже). Кломифен-цитрат (кломид) отличается особенно высоким сродством к рецепторам эстрогенов в гипоталамусе. Кломифен вначале предполагали использовать как противозачаточное средство, но оказалось, что он обладает противоположным действием. Поскольку кломифен конкурирует с эстрадиолом за гипоталамические рецепторы, выделение гонадолиберина перестает ограничиваться и высвобождаются большие количества ЛГ и ФСГ. Под действием кломифена часто наблюдается одновременное созревание многих фолликулов, что может приводить к многоплодной беременности. Нафоксидин (нестероидное соединение) и тамоксифен взаимодействуют с рецепторами эстрогенов, образуя с хроматином очень стабильный комплекс. Поскольку связанные таким образом рецепторы не могут вступать в новый цикл, эти соединения надолго ингибируют действие эстрадиола. Такие антагонисты используют для лечения эстроген-зависимого рака молочной железы.
Б. Протестины. Синтез соединений, которые, обладая прогестиновой активностью, не оказывали бы при этом эстрогенного или андрогенного действия, оказался делом нелегким. 17а-Алкилзамещенные 19-нортестостерона (например, норэтиндрон) характеризуются минимальной андрогенной активностью у большинства женщин и используются в качестве пероральных контрацептивов. Другой активный прогестин — медроксипрогестерон-ацетат. Внутримышечное введение медроксипрогестерона пролонгированного действия приводит к подавлению овуляции на несколько месяцев. Однако гораздо чаще этот препарат используется для лечения дифференцированного рака эндометрия. Считают, что он блокирует деление нормальных и злокачественных клеток эндометрия путем образования стабильных комплексов с рецепторами прогестерона (что препятствует действию природного гормона).
