Биохимия человека Том 2 - Марри Р. 1993
Частные вопросы
Сократительные и структурные белки
Мышцы
Мышцы — главный биохимический преобразователь потенциальной (химической) энергии в кинетическую (механическую). Мышечная ткань занимает первое место по объему среди других тканей человека; на ее долю при рождении приходится чуть меньше 25%, у людей среднего возраста — более 40%, а у пожилых — чуть меньше 30% общей массы тела.
Эффективное преобразование химической энергии в механическую возможно при соблюдении ряда условий: 1) должно быть обеспечено постоянное снабжение химической энергией. В мышцах позвоночных химическая энергия заключена в молекулах АТР и креатинфосфата; 2) должны существовать средства регуляции механической активности, т.е. в случае мышц — скорости, длительности и силы сокращения; 3) процесс преобразования должен находиться под контролем оператора, в данном случае его функцию выполняет нервная система; 4) для того чтобы «машина» преобразования энергии могла использоваться многократно, необходим механизм возврата системы в исходное состояние.
Мышцу можно сравнивать с машиной, которая «тянет», но не «толкает», следовательно, каждая мышца должна находиться под антагонистическим воздействием другой группы мышц или какой-либо иной силы, такой, как сила тяжести или эластичная отдача.
В организме позвоночных существуют три типа мышц: скелетные, сердечные и гладкие. Скелетные и сердечные мышцы при микроскопическом исследовании обнаруживают поперечную исчерченность; в гладких мышцах такая исчерченность отсутствует. В то время как скелетные мышцы находятся под волевым нервным контролем, сердечная и гладкая мышцы функционируют непроизвольно.
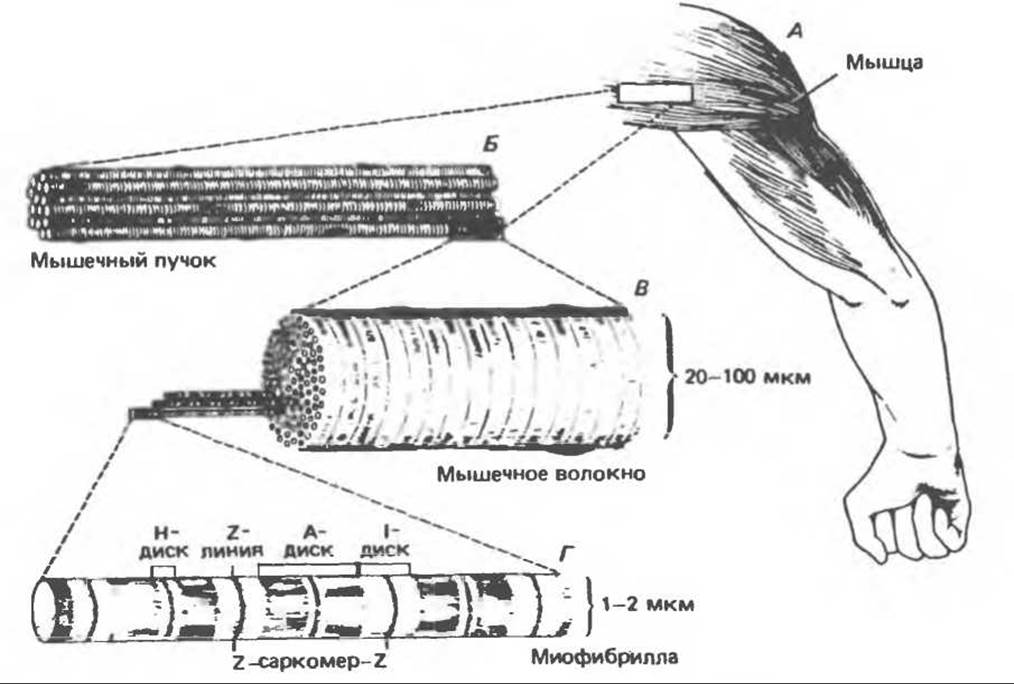
Рис. 56.1. Структура поперечнополосатой мышцы. (Drawing by Sylvia Colard Keene. Reproduced, with permission, from Bloom W., Fawcett D. W. A Textbook of Histology, 10th ed. Saunders, 1975.)
Структура мышц
Поперечнополосатая мышца состоит из многоядерных клеток (мышечных волокон), окруженных электровозбудимой мембраной — сарколеммой. При микроскопическом исследовании отдельной мышечной клетки, которая может быть вытянута во всю длину мышцы, в ней обнаруживается пучок, состоящий из множества параллельно расположенных миофибрилл; они погружены во внутриклеточную жидкость, называемую саркоплазмой. Эта жидкость содержит гликоген, макроэргические соединения (АТР и фосфокреатин) и ферменты гликолиза.
Саркомер — это функциональная единица мышцы. Саркомеры следуют друг за другом вдоль оси фибриллы, повторяясь через каждые 1500—2300 нм (рис. 56.1). При исследовании миофибриллы в электронном микроскопе выявляется чередование темных и светлых дисков (диски А и 1). Центральная область диска А (зона Н) выглядит менее плотной, чем остальная его часть. Диск I делит пополам очень плотная и узкая линия Z. Эти детали мышечной структуры представлены на рис. 56.2.
Исчерченность мышц, видимая под световым микроскопом, — это результат высокой степени их организации, когда большинство мышечных клеток выстраивается таким образом, что их саркомеры располагаются параллельно (рис. 56.1).
Исследование поперечных срезов миофибрилл в электронном микроскопе показывает, что каждая миофибрилла состоит из двух типов продольных филаментов (нитей). Первый тип («толстые» нити)
ограничены A-диском, они состоят главным образом из белка миозина, имеют около 16 нм в диаметре и образуют на поперечном срезе шестиугольник (рис. 56.2). Второй тип филаментов («тонкие» нити) занимает I-диск, распространяется на диск А, но не достигает его Н-зоны (рис. 56.2). Диаметр тонких нитей составляет около 6 нм. Они содержат белки актин, тропомиозин и тропонин. В диске А тонкие нити располагаются вокруг толстого (миозинового) филамента в виде второго шестиугольника. Таким образом, каждый тонкий филамент занимает симметричное положение между тремя толстыми филаментами, а каждый толстый филамент симметрично окружен шестью тонкими филаментами (рис. 56.2).
Толстые и тонкие филаменты взаимодействуют при посредстве поперечных мостиков, расположенных вдоль толстого филамента с промежутками в 14 нм. Как показано на рис. 56.2, поперечные мостики, или «наконечники», толстых филаментов имеют противоположные полярности на двух концах филамента. Эти полярные концы разделены сегментом (полосой М) длиной 150 нм, не содержащим выростов.
Во время сокращения мышцы длина толстых и тонких филаментов не меняется, но Н-зона и 1-диски укорачиваются, следовательно, переплетающиеся филаменты должны скользить относительно друг друга. Напряжение, развивающееся при сокращении мышцы, пропорционально степени перекрывания филаментов и, следовательно, числу поперечных мостиков. «Головка» каждого поперечного мостика соединена с толстым филаментом гибким волокнистым сегментом, который может изгибаться, регулируя пространство между филаментами.
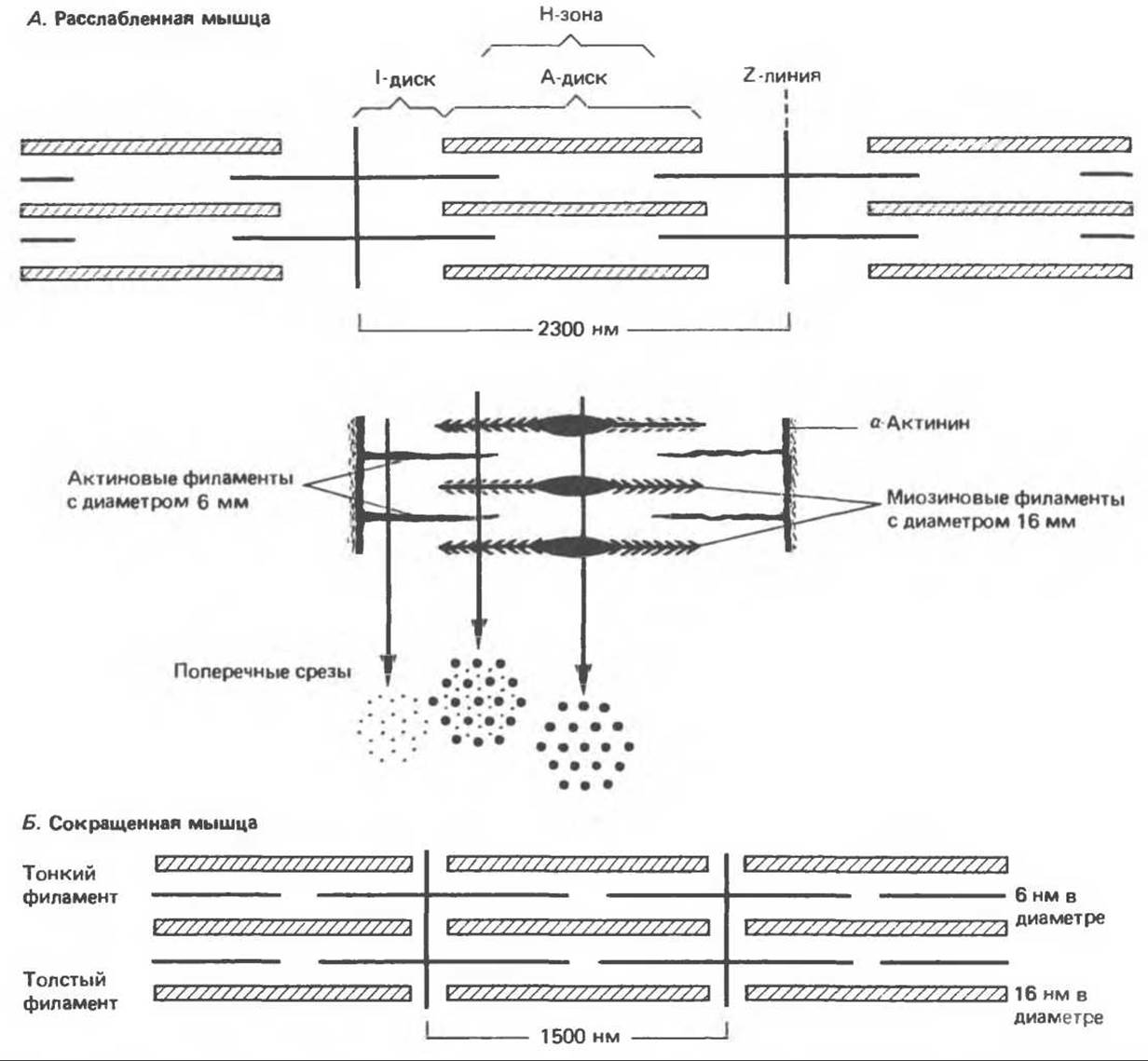
Рис. 56.2. Расположение филаментов в поперечнополосатой мышце. А. Расслабленная мышца. Б. Сокращенная мышца.
Мышечные белки
Масса свежих мышечных волокон на 75% состоит из воды и содержит более 20% белка. Два главных мышечных белка — актин и миозин.
Мономерный (глобулярный) актин (G-актин) — это глобулярный белок с мол. массой 43000, на долю которого приходится 25% общей массы мышечного белка. При физиологической величине ионной силы и в присутствии магния G-актин подвергается нековалентной полимеризации с образованием нерастворимого двойного спирального филамента, получившего название F-актин (рис. 56.3). Волокно F-актина имеет толщину 6—7 нм и через каждые 35,5 нм — повторяющиеся структурные элементы. Ни G-, ни F-актин не обладают каталитической активностью.
В поперечнополосатых мышцах присутствуют еще четыре белка, минорных в плане их вклада в массу мышечной ткани, но выполняющих важную функцию. Тропомиозин представляет собой вытянутую в виде тяжа молекулу, состоящую из двух цепей, а и ß, и примыкающую к F-актину в щели между двумя полимерами (рис. 56.3). Этот белок имеется во всех мышцах и подобных им структурах. Характерной особенностью именно поперечнополосатых мышц является наличие в них тропониновой системы, включающей три разных белка. Тропонин Т (ТnТ) так же, как и два других тропониновых компонента, связывается с тропомиозином (рис. 56.3). Тропонин I (TnI) ингибирует взаимодействие между F-актином и миозином и также связывается с другими компонентами тропонина. Тропонин С (ТnС) — кальций-связывающий белок, первичная и вторичная структура, а также функция которого, совершенно аналогичны соответствующим характеристикам широко распространенного в природе белка — кальмодулина. И тропонин С, и кальмодулин связывают четыре иона кальция на молекулу белка и имеют мол. массу 17000. Тонкий филамент поперечнополосатой мышцы состоит из F-актина, тропомиозина и трех тропониновых компонентов: ТnС, TnI и ТnТ (рис. 56.3). Тропомиозин и тропониновая система чередуются через каждые 38,5 нм.
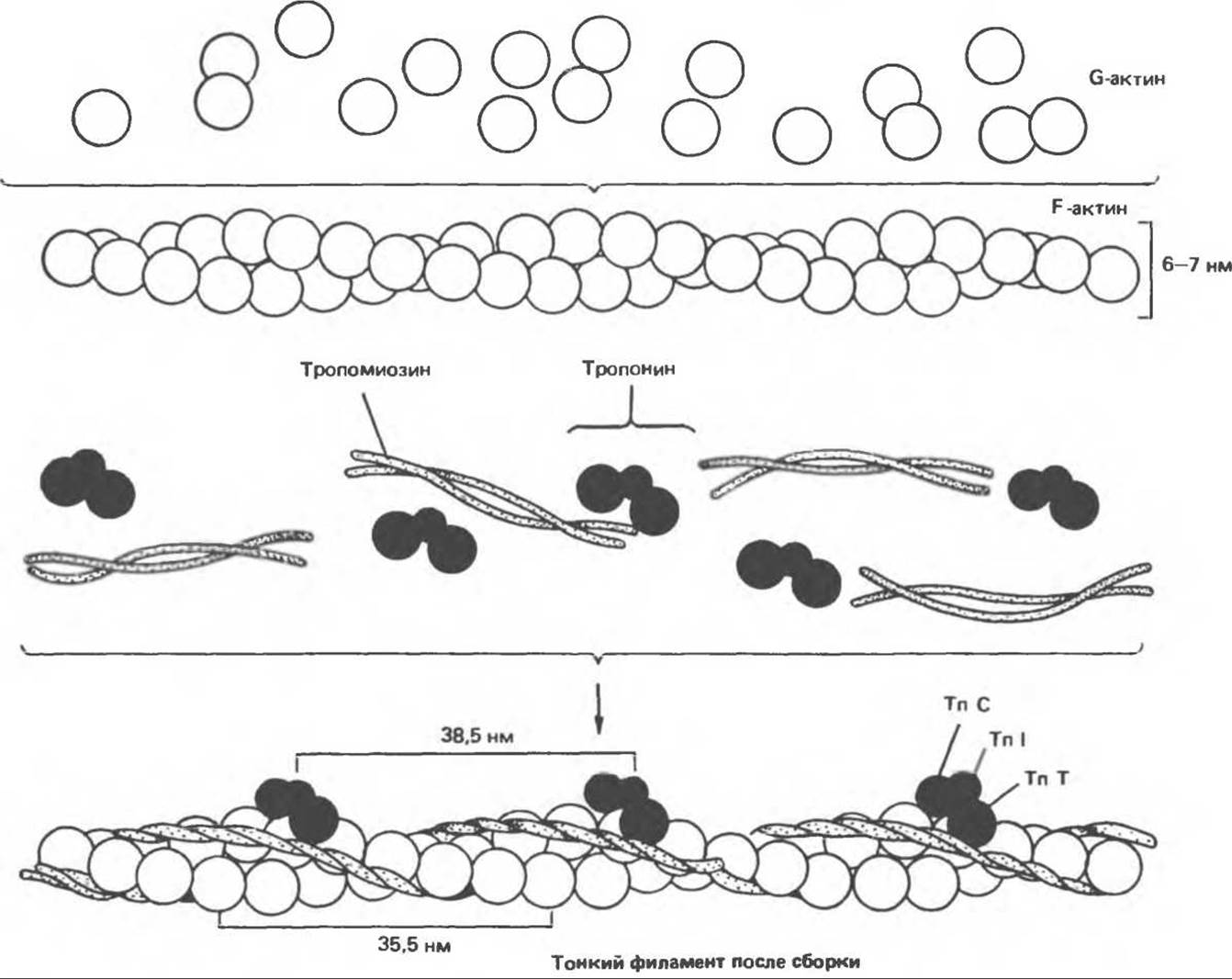
Рис. 56.3. Схематическое изображение тонкого филамента. Показана пространственная конфигурация трех главных белковых компонентов: актина, тропомиозина и тропонина.
Миозин по массе составляет 55% мышечного белка и образует толстые филаменты (нити). Он представляет собой асимметричный гексамер с мол. массой 460000. В миозине различают фибриллярную часть, состоящую из двух переплетенных спиралей, каждая из которых имеет на одном конце глобулярную «головку» (рис. 56.4). Гексамер включает одну пару тяжелых цепей (мол. масса 200 000) и две пары легких цепей (мол. масса 15000—27000). Миозин скелетных мышц обладает АТР-гидролизующей (АТР-азной) активностью и связывается с нерастворимой молекулой — F-актином.
Большая часть сведений о миозине получена при изучении продуктов его частичного гидролиза. Обработка миозина трипсином приводит к образованию двух фрагментов — меромиозинов. Легкий меромиозин (ЛММ) состоит из агрегированных нерастворимых а-спиральных фибрилл (рис. 56.5). Он не обладает АТРазной активностью и не связывается с F-актином.
Тяжелый меромиозин (ТММ) представляет собой растворимый белок с мол. массой 340 000, содержащий и фибриллярный, и глобулярный фрагменты (рис. 56.5). Он обладает АТРазной активностью и связывается с F-актином. При гидролизе ТММ папаином образуются два субфрагмента, S-1 и S-2. S-2 имеет фибриллярную структуру, не проявляет АТР- азной активности и не связывает F-актин.

Рис. 56.4. Схема молекулы миозина с двумя переплетенными а-спиралями (фибриллярная часть), глобулярной областью (G) и легкими цепями (L).
S-1 характеризуется мол. массой 115000, проявляет АТРазную активность и в отсутствие АТР связывает актин, снабжая его «наконечниками» (рис. 56.6). Хотя и S-1, и ТММ сами обладают АТРазной активностью, но при добавлении F-актина эта активность возрастает в 100—200 раз. Показано, что F-актин резко ускоряет освобождение продуктов действия миозиновой АТРазы — ADP и неорганического фосфата. Таким образом, хотя F-актин сам по себе не влияет на гидролиз АТР, его способность стимулировать освобождение продуктов АТРазной реакции обеспечивает значительное увеличение общей скорости катализа.
а-Актинин — это обнаруживаемая в зоне Z-линии белковая молекула, к которой присоединяются концы F-актиновых молекул тонких филаментов (рис. 56.2).
Молекулярная функция мышц
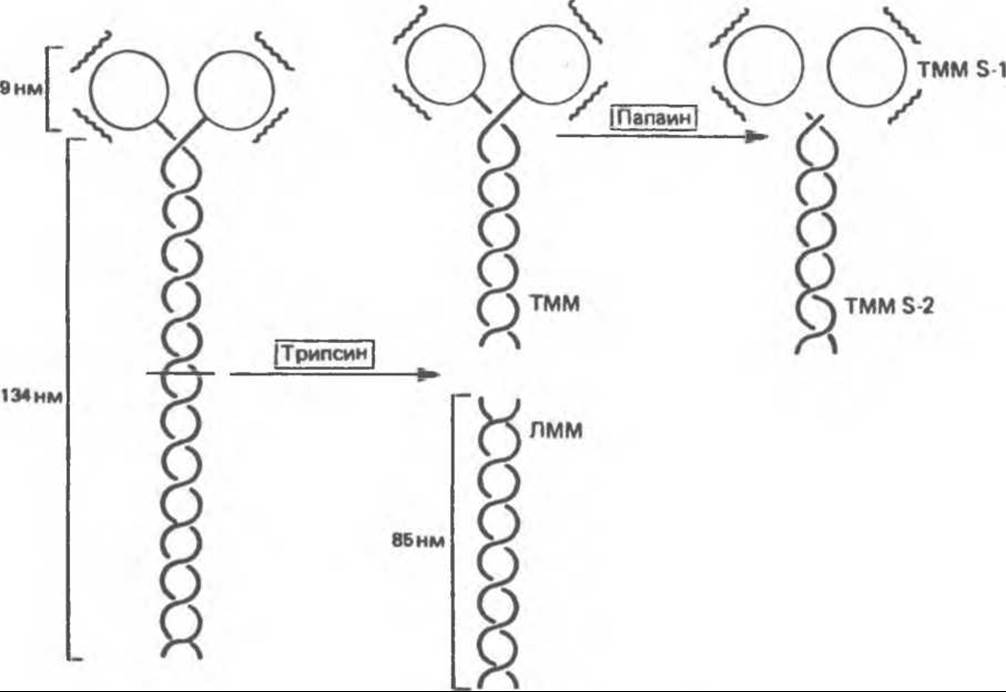
Рис. 56.5. Ферментативное расщепление миозина. ТММ — тяжелый меромиозин; ЛММ — легкий меромиозин; S-1 — фрагмент 1; S-2—фрагмент 2.
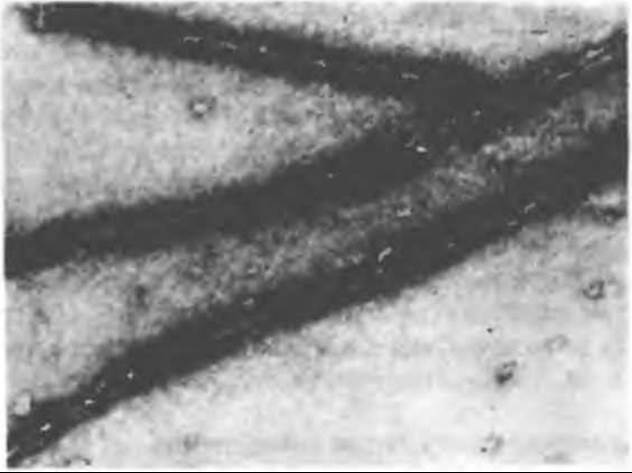
Рис. 56.6. Образование «наконечников» при связывании актиновых филаментов с S-1-фрагментами миозина. (Courtesy of Professor James Spudich, Stanford University.)
Вопрос о связи структуры и функции мышц может быть сформулирован в биохимических терминах следующим образом: каким образом гидролиз АТР приводит к видимому невооруженным глазом движению? Как отмечалось выше, мышечное сокращение состоит из циклов присоединения и отсоединения глобулярной «головки» миозина от нити F-актина. Присоединение сопровождается изменением актин — миозинового взаимодействия, так что актиновые и миозиновые филаменты скользят относительно друг друга. Энергия для этого скольжения поставляется за счет гидролиза АТР. Гидролиз АТР миозиновой АТРазой значительно ускоряется при связывании миозиновой «головки» с F-актином. Биохимический цикл мышечного сокращения состоит из пяти стадий (рис. 56.7).
1) Миозиновая головка сама по себе может гидролизовать АТР до ADP и неорганического фосфата, но не обеспечивает освобождение продуктов гидролиза. Следовательно, этот процесс носит скорее стехиометрический, чем каталитический характер.
2) Миозиновая головка, содержащая ADP и неорганический фосфат, может свободно вращаться под большими углами и (при достижении нужного положения) связываться с F-актином, образуя с осью фибриллы угол около 90°.

Рис. 56.7. Г идролиз АТР запускает цикл ассоциации и диссоциации актина и миозина в пяти реакциях, описанных в тексте; Pi — неорганический фосфат. (Modified from Stryer L. Biochemistry, 2nd ed. Freeman, 1981.)
3) Это взаимодействие обеспечивает высвобождение ADP и неорганического фосфата из актин — миозинового комплекса. Поскольку наименьшую энергию актомиозиновая связь имеет при величине угла 45°, миозин изменяет свой угол с осью фибриллы с 90° на примерно 45°, продвигая актин (на 10—15 нм) в направлении центра саркомера.
4) Новая молекула АТР связывается с комплексом миозин — F-актин.
5) Комплекс миозин — АТР обладает низким сродством к актину и поэтому происходит отделение миозиновой (АТР) головки от F-актина. Последняя стадия и есть собственно расслабление, которое таким образом отчетливо зависит от связывания АТР с актин — миозиновым комплексом. АТР вновь гидролизуется миозиновой головкой без высвобождения ADP и неорганического фосфата, и цикл возобновляется.
Таким образом, АТР отсоединяет миозиновую головку от тонкой нити и является движущей силой сокращения. Эффективность такого сокращения — около 50%; эффективность двигателя внутреннего сгорания — менее 20%.
Регуляция сокращения и расслабления мышц
Сокращение любых мышц протекает по общему механизму, описанному выше. Мышечные волокна разных организмов и даже разных тканей одного организма могут обладать различными молекулярными механизмами регуляции их сокращения и расслабления. Заметим, что во всех случаях ключевая регуляторная роль принадлежит ионам Са2+. Существуют два главных механизма регуляции мышечного сокращения: актиновый и миозиновый.
Актиновая регуляция
Актиновая регуляция характерна для поперечнополосатых мышц позвоночных — скелетных и сердечной. Согласно общему механизму, рассмотренному выше, единственным потенциально лимитирующим фактором в цикле мышечного сокращения может быть АТР. Скелетные мышцы ингибируются в покое и деингибируются с активацией сокращения. Роль ингибитора в поперечнополосатых мышцах выполняет тропониновая система, связанная в тонких филаментах с тропомиозином и F-актином (рис. 56.3). При отсутствии тропомиозин-тропониновой системы регуляция сокращения поперечнополосатых мышц (или АТРазы как биохимического индикатора сокращения) не осуществляется. Как отмечалось выше, тропомиозин локализуется в щели F-актина, а три компонента тропонина — ТnТ, ТnІ и ТnС — связаны с комплексом F-актин — гропомиозин. TnI предотвращает присоединение миозиновой головки к соответствующему связывающему сайту F-актина, либо изменяя конформацию F-актина (через тропо- миозиновые молекулы), либо просто перемещая («вращая») тропомиозин в то положение, в котором он блокирует сайты связывания миозиновых головок на F-актине. В любом случае предотвращается активация миозиновой АТРазы, которая опосредована этим связыванием. Таким образом, система TnІ блокирует цикл сокращения на стадии 2 схемы, представленной на рис. 56.7. Именно это лежит в основе ингибированного состояния расслабленной поперечнополосатой мышцы.
Мышечное сокращение опосредуется Са2+. В саркоплазме покоящейся мышцы концентрация кальция составляет 10-7—10-8 моль/л. Кальций попадает в саркоплазматический ретикулум в результате активного транспорта при участии Са-связывающего белка, называемого кальсеквестрином. Саркомер окружен возбудимой мембраной с поперечными каналами, подходящими к саркоплазматическому ретикулуму. При возбуждении мембраны саркомера, например, в случае взаимодействия ацетилхолиновых рецепторов с ацетилхолином, Са2+ быстро высвобождается из саркоплазматического ретикулума в саркоплазму, так что его концентрация в ней возрастает до 10-5 моль/л. Са2+-связывающие сайты на ТnС в тонком филаменте быстро насыщаются Са2+. Комплекс ТnС-4Са2+ реагирует с Tnl и ТnТ, влияя на их взаимодействие с тропомиозином. Последний в соответствии с этим либо просто отсоединяется, либо изменяет конформацию F-актина таким образом, что появляется возможность взаимодействия ADP-Рi-миозиновой головки с F-актином и начинается сократительный цикл.
Расслабление происходит, когда 1) содержание Са2+ в саркоплазме падает ниже 10-7 моль/л вследствие его поглощения саркоплазматическим ретикулумом; 2) комплекс ТnС-4Са2+ утрачивает свой Са2+; 3) тропонин, реагируя с тропомиозином, ингибирует дальнейшее взаимодействие миозиновой головки с F-актином и 4) миозиновые головки в присутствии АТР отделяются от F-актина, вызывая расслабление. Таким образом, Са2+ регулирует мышечное сокращение при помощи аллостерического механизма, опосредованного в мышце ТnС, Tnl, ТnТ, тропомиозином и F-актином.
В сердечной мышце основным источником ионов Са2+ для возбуждения служит внеклеточная жидкость. Если Са2+ во внеклеточной жидкости отсутствует, сокращения сердечной мышцы прекращаются в течение одной минуты; скелетная мышца в таких условиях может сокращаться часами.
Исчезновение АТР из саркоплазмы приводит к следующим последствиям; 1) Са2+-насос саркоплазматического ретикулума перестает поддерживать низкую концентрацию Са2+ в саркоплазме; при этом стимулируется взаимодействие миозиновых головок с F-актином; 2) не происходит зависимого от АТР отделения миозиновых головок от F-актина, при этом наступает трупное окоченение.
Мышечное сокращение не принадлежит к ряду феноменов «все или ничего», как может показаться читателю. Оно представляет собой тонкое динамическое равновесие между процессами присоединения и отделения миозиновых головок от F-актина. Система находится под сложным регуляторным влиянием со стороны нервной системы.
Миозиновая регуляция сокращения
Как отмечалось выше, во всех мышцах присутствуют актин, миозин и тропомиозин, но тропониновую систему содержат только поперечнополосатые мышцы позвоночных. Следовательно, механизмы регуляции сокращения в разных сократительных системах должны различаться.
Молекулярные структуры гладких мышц весьма сходны с соответствующими структурами поперечнополосатых мышц, но расположение саркомеров в них не дает характерную для поперечнополосатых мышц картину исчерченности. Подобно скелетным мышцам, гладкие мышцы содержат молекулы а-актинина и тропомиозина, но не обладают тропониновой системой; кроме того, легкие цепи миозиновых молекул гладких мышц отличаются от аналогичных цепей поперечнополосатых мышц. Тем не менее сокращение гладких мышц, как и сокращение поперечнополосатых, регулируется Са2+.
Когда миозин гладких мышц связывается с F-актином в отсутствие других мышечных белков, таких, как тропомиозин, образующийся комплекс лишен заметной АТРазной активности. Это резко отличается от ситуации, характерной для взаимодействия с F-актином миозина поперечнополосатых мышц, когда регистрируется высокая активность АТРазы. Миозин гладкой мускулатуры содержит легкую цепь (p-легкую цепь), предотвращающую связывание миозиновых головок с F-актином. Для того чтобы эта легкая цепь не препятствовала активации миозиновой АТРазы при взаимодействии с F- актином, она должна предварительно подвергнуться фосфорилированию. Фосфорилирование легкой цепи р запускает процессы ассоциации — диссоциации в сократительном цикле гладкой мускулатуры.
В саркоплазме гладких мышц присутствует киназа легких цепей миозина, зависимая от кальция. Для активации этого фермента кальцием требуется связывание его субъединицы, имеющей молекулярную массу 105000, с кальмодулином∙4Са2+ (рис. 56.8). Активированная кальмодулином∙4Са2+ киназа легких цепей фосфорилирует легкую цепь р, которая при этом перестает ингибировать взаимодействие миозина c F-актином. Таким образом, начинается сократительный цикл (рис. 56.8).

Рис. 56.8. Кальциевая регуляция сокращения гладких мышц. (Adapted from Adelstein R. S., Eisenberg R. Regulation and kinetics of actin myosin ATP interaction. Annu. Rev. Biochem., 1980, 49, 921.)
Расслабление гладких мышц происходит, когда 1) содержание ионов Са2+ в саркоплазме падает ниже 10-7 моль/л; 2) Са2+ отсоединяется от кальмодулина, который в свою очередь отделяется от киназы легкой цепи миозина, вызывая ее инактивацию 3) нового фосфорилирования легкой цепи р не происходит, и протеин фосфатаза легкой цепи, которая постоянно активна и не зависит от кальция, отщепляет от легкой цепи р ранее присоединившиеся к ней фосфаты; 4) дефосфорилированная легкая цепь р миозина ингибирует связывание миозиновых головок с F-актином и подавляет активность АТРазы; 5) миозиновые головки в присутствии АТР отделяются от F-актина, а повторное их связывание произойти не может из-за присутствия в системе дефосфорилированной легкой цепи р. В результате описанных событий происходит расслабление мышцы.
В табл. 56.1 суммируются и сравниваются данные о регуляции актин-миозинового взаимодействия (активации миозиновой АТРазы) в поперечнополосатых и гладких мышцах.
Киназа легких цепей миозина не является прямым объектом активации со стороны сАМР. Тем не менее обычная активируемая сАМР протеинкиназа (см. гл. 44) может фосфорилировать этот фермент (не легкую цепь р саму по себе). Фосфорилированная киназа легких цепей миозина обладает значительно меньшим сродством к кальмодулин Са2+ и потому менее чувствительна к активации. Соответственно повышение уровня сАМР уменьшает сократительную реакцию гладких мышц на увеличение содержания Са2+ в саркоплазме. Описанный молекулярный механизм может объяснить расслабляющее действие на гладкие мышцы ß-адренергической стимуляции. Фенотиазины широко применяющиеся в качестве антипсихотических средств, связываются с кальмодулином и предотвращают его взаимодействие с кальций-зависимыми ферментами. Фенотиазины вызывают также расслабление гладкой мускулатуры.
Поперечнополосатые мышцы моллюсков, таких, как морской гребешок, обладают системой миозиновой регуляции сокращений. Миозин и F актин гребешка, подобно этим белкам в гладких мышцах, лишены АТРазной активности, что обусловлено ингибиторными свойствами «регуляторной» легкой цепи миозина гребешка. Торможение актин- миозинового взаимодействия у морского гребешка снимается при прямом связывании Са2+ со специфическим центром молекулы миозина. Этот регуляторный механизм не требует ковалентной модификации миозина и (или) добавления специфических белков, таких, как кальмодулин или ТnС.
Таблица 56.1. Актин-миозиновые взаимодействия в поперечнополосатых и гладких мышцах
|
Поперечнополосатые мышцы |
Гладкие мышцы (и немышечные клетки) |
|
|
Белки мышечных филаментов |
Актин |
Актин |
|
Миозин (гексамер) |
Миозин (гексамер) 1) |
|
|
Тропомиозин |
Тропомиозин |
|
|
Тропонин (TnI, ТnТ, ТnС) |
||
|
Спонтанное взаимодействие F-актина с одним миозином (спонтанная активация миозиновой АТРазы F-актином) |
Есть |
Нет |
|
Ингибитор взаимодействия F-актина с миозином (ингибитор F-актин-зависимой активации АТРазы) |
Тропониновая система (TnI) |
Нефосфорилированная р-легкая цепь миозина |
|
Сокращение активируется |
Са2+ |
Са2+ |
|
Прямое действие Са2+ |
4Са2+ связываются с ТnС |
4Са2+ связываются с кальмодулином |
|
Действие связанного с белком Са2+ |
ТnС∙4Са2+ препятствует ингибирующему эффекту TnI на взаимодействие F-актина с миозином (делает возможной активацию АТРазы F-актином) |
Кальмодулин-4Са2+ активирует киназу легких цепей миозина, которая фосфорилирует p-легкую цепь миозина. Фосфорилированная р-легкая цепь перестает ингибировать взаимодействие F-актина с миозином (делает возможной активацию АТРазы F-актином) |
1) Легкие цепи миозина в поперечнополосатых и гладких мышцах различаются.
Фосфорилирование мышечных белков
Фосфорилирование легкой цепи миозина гладких мышц снимает ее ингибиторное влияние на взаимодействие актина с миозином и тем самым запускает сократительный цикл. Таким образом, для начала взаимодействия актина с миозином в гладких мышцах требуется фосфорилирование.
Одна из пар легких цепей миозина скелетных мышц также может подвергаться фосфорилированию, которое, однако, не влияет на активируемую актином миозиновую АТРазу (что характерно для миозина гладких мышц). Предполагается, что фосфат на легких цепях миозина может образовывать хелат с Са2+ (связанным с комплексом тропомиозин-TnC-актин), увеличивая тем самым скорость образования поперечных мостиков между миозиновыми головками и актином.
Некоторые новые данные свидетельствуют о том, что фосфорилирование тяжелых цепей миозина служит необходимым условием для их сборки в толстые филаменты в скелетных мышцах, гладких мышцах и немышечных клетках (см. ниже).
TnI и пептидный компонент Са2+-насоса саркоплазматического ретикулума в сердечной мышце могут фосфорилироваться сАМР-зависимой протеинкиназой. Между фосфорилированием TnI и усилением сокращений сердечной мышцы, вызываемым катехоламинами, имеется некоторая корреляция. Этот механизм может обусловливать инотропный эффект (повышение сократимости) ß-адренергических соединений на сердце.
Метаболизм мышц
АТР, необходимый в качестве постоянного источника энергии для мышечного цикла сокращение — расслабление, может образовываться за счет гликолиза, окислительного фосфорилирования, креатинфосфата или двух молекул ADP. Запасы АТР в скелетной мышце при сокращении быстро истощаются, и их хватает менее чем на секундное сокращение. В медленных скелетных мышцах, обладающих значительными резервами O2 в миоглобине, основной источник регенерации АТР — окислительное фосфорилирование. Быстрые скелетные мышцы регенерируют АТР главным образом в ходе гликолиза.
Фосфагены, такие как креатинфосфат, предотвращают быстрое истощение запасов АТР, поставляя легко используемый макроэргический фосфат, необходимый для ресинтеза АТР из ADP. Креатинфосфат образуется из АТР и креатина в период расслабления мышцы, когда потребность в АТР не столь велика. Фосфорилирование креатина катализируется креатинфосфокиназой (КФК) — специфичным для мышц ферментом, который используется при диагностике осгрых или хронических мышечных нарушений.
Саркоплазма скелетных мышц содержит большие запасы гликогена, локализованного в гранулах, примыкающих к 1-диску. Высвобождение глюкозы из гликогена зависит от специфического мышечного фермента гликогенфосфорилазы (см. гл. 19). Для того чтобы обеспечить образование глюкозо-6-фосфата, используемого в процессе гликолиза, гликогенфосфорилаза b должна быть активирована в фосфорилазу а. Для активации требуется фосфорилирование фосфорилазы b, осуществляемое фосфорилаза-b-киназой (см. гл. 19). Активация фосфорилаза-b-киназы, которая также осуществляется путем фосфорилирования фермента, стимулируется Са2+. Таким образом, Са2+ не только стимулирует мышечное сокращение, но и усиливает образование необходимого для этого процесса источника энергии — АТР. Мышечная гликогенфосфорилаза b отсутствует при специфическом заболевании мышц (болезнь Мак-Ардля), которое представляет собой одну из форм гликогенозов.
АТР в мышечной ткани образуется и в ходе окислительного фосфорилирования — процесса, требующего постоянного притока кислорода. Мышцы, характеризующиеся высокой потребностью в кислороде в связи с длительным состоянием сокращения (например, для поддержания определенной позы), обладают способностью резервировать кислород в миоглобине (см. гл. 6). Поскольку кислород связывается в миоглобине с гемом, мышцы, содержащие миоглобин, окрашены в красный цвет в отличие от не содержащих его белых скелетных мышц. В табл. 56.2 приведены сравнительные данные о некоторых свойствах быстрых (или белых) и медленных (или красных) скелетных мышц.
Миоаденилаткиназа — фермент, присутствующий в мышцах, катализирует образование одной молекулы АТР и одной молекулы АМР из двух молекул ADP. Эта реакция, показанная на рис. 56.9, сопряжена с гидролизом АТР миозиновой АТРазой во время мышечного сокращения. На рисунке, кроме того, отражена связь между различными источниками АТР и его потреблением в ходе сокращения мышцы.
У людей главным после жира источником запасенной энергии служит белок скелетных мышц. Это объясняет очень большую потерю мышечной массы (особенно у взрослых людей), наблюдающуюся при длительной калорической недостаточности.
Изучению распада тканевого белка in vivo препятствует тот факт, что аминокислоты, высвобождающиеся при внутриклеточной деградации белков, могут в значительной степени реутилизироваться для синтеза белка в клетке или переноситься к другим органам и вступать там в анаболические процессы. Однако актин и миозин после синтеза их пептидных связей метилируются с образованием 3-метилгистидина. В ходе внутриклеточного распада актина и миозина 3-метилгистидин высвобождается и выделяется с мочой. При введении метки крысам или человеку было показано, что экскреция с мочой метилированной аминокислоты служит надежным показателем скорости деградации белка миофибрилл в мышцах. Фракционная скорость распада мышечного белка у пожилых людей мало отличается от этого показателя у молодых, но, поскольку масса мышц при старении уменьшается, снижается и вклад этой ткани в общее возрастное увеличение распада белков в организме.
Таблица 56.2. Характеристика быстрых и медленных скелетных мышц
|
Быстрая скелетная мышца |
Медленная скелетная мышца |
|
|
Активность миозиновой АТРазы |
Высокая |
Низкая |
|
Утилизация энергии |
Высокая |
Низкая |
|
Цвет |
Белый |
Красный |
|
Миоглобин |
Нет |
Есть |
|
Частота сокращений |
Высокая |
Низкая |
|
Длительность сокращений |
Малая |
Большая |
Как отмечалось выше, скелетные мышцы служат основным резервом белка в организме. Они обладают также высокой активностью в отношении деградации одних и синтеза других аминокислот. У млекопитающих именно мышцы являются главным местом катаболизма аминокислот с разветвленной цепью. Мышечная ткань окисляет лейцин до СО2 и превращает углеродный скелет аспартата, аспарагина, глутамата, изолейцина и валина в интермедиаты цикла трикарбоновых кислот. Способность мышц разрушать аминокислоты с разветвленной цепью при голодании и диабете возрастает в 3— 5 раз.
Мышцы также синтезируют и высвобождают большие количества аланина и глутамина. В синтезе этих соединений используются аминогруппы, которые образуются при распаде аминокислот с разветвленной цепью и затем переносятся на а-кетоглутарат и пируват в ходе реакций трансаминирования. Источником почти всего пирувата, идущего на синтез аланина, является гликолиз из экзогенной глюкозы. Эти реакции формируют так называемый глюкозо-аланиновый цикл, в котором аланин мышц используется в процессе печеночного глюконеогенеза и в то же время доставляет в печень аминогруппы, удаляемые в виде мочевины.
Углеродный скелет аминокислот, подвергшихся деградации и включившихся в цикл трикарбоновых кислот в мышечной ткани, превращается главным образом в глутамин и пируват, который далее окисляется или превращается в лактат. Таким образом, при голодании или в период после всасывания большая часть образующихся в процессе распада мышечного белка аминокислот покидает мышцы; исключением являются изолейцин, валин, глутамат, аспартат и аспарагин: они участвуют в образовании глутамина, который высвобождается мышцами и используется другими тканями.
Давно известно, что работающая мышца высвобождает аммиак. Как установлено в настоящее время, непосредственным источником аммиака в скелетной мышце служит АМР, который дезаминируется в IMP под действием аденилатдезаминазы. IMP может вновь превращаться в АМР в ходе реакций, использующих аспартат и катализируемых аденилсукцинатсинтетазой и аденилсукциназой (см. гл. 35).

Рис. 56.9. Множественные источники АТР в мышце.