Биохимия человека Том 2 - Марри Р. 1993
Биохимия внутри- и межклеточных коммуникаций
Мембраны: структура, сборка и функции
Жидкостно-мозаичная модель мембран
Функционирующие мембраны представляют собой двумерный раствор глобулярных интегральных белков, диспергированных в жидком фосфолипидном матриксе. Жидкостно-мозаичная модель мембранной структуры была предложена в 1972 г. Сингером и Николсоном (рис. 42.9). Первые данные об адекватности этой модели были получены при искусственно индуцированном слиянии двух разных родительских клеток. Оказалось, что при образовании межвидовой гибридной клетки в плазматической мембране происходит быстрое стохастическое перераспределение видоспецифичных белков. Впоследствии было показано, что фосфолипиды тоже способны быстро перераспределяться в плоскости мембраны. Такая диффузия в плоскости мембраны, называемая латеральной, может осуществляться довольно быстро; одна молекула фосфолипида перемещается за 1 с на расстояние несколько микрометров.
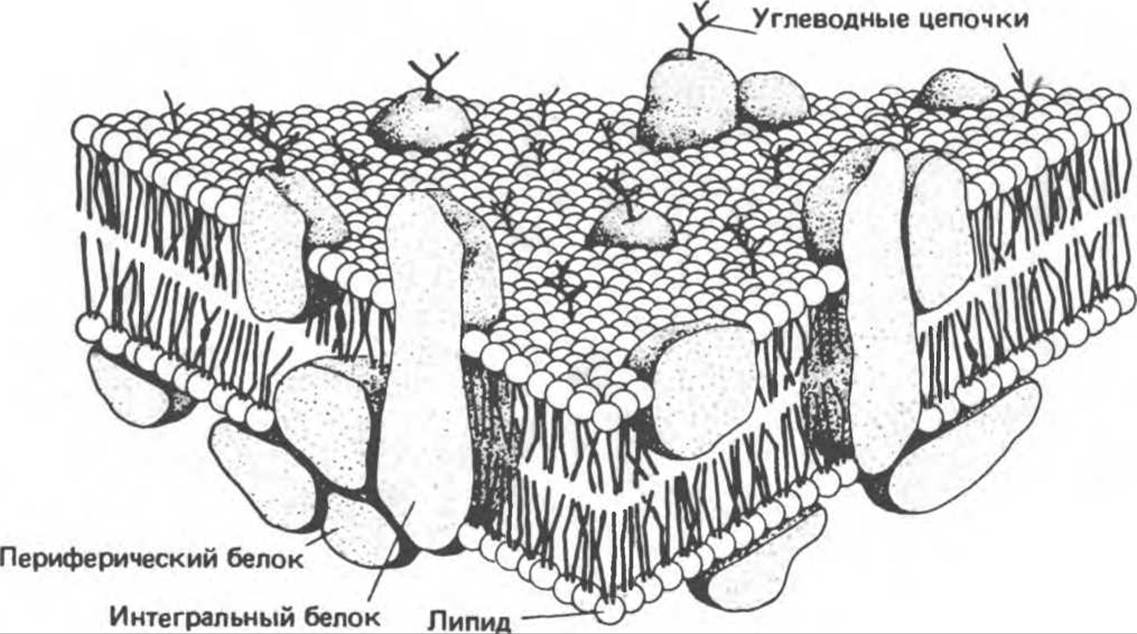
Рис. 42.9. Жидкостно-мозаичная модель мембранной структуры. Основой мембраны является липидный бислой; с ним связаны белки, либо погруженные в бислой, либо присоединенные к цитоплазматической поверхности. Интегральные мембранные белки жестко закреплены в липидном бислое. Некоторые из этих белков пронизывают бислой и называются трансмембранными, другие погружены либо в наружный, либо во внутренний слой. Белки, слабо связанные с внутренней поверхностью мембраны, называются периферическими. Многие белки и липиды несут олигосахаридные цепочки, выступающие во внешнюю среду. (Из работы Junqueira L. С., Carneiro J., Long J. A.: Basic Histology, 5th ed. Appleton and Lange. 1986, с любезного разрешения.)
Фазовые переходы и, следовательно, текучесть мембран сильно зависят от липидного состава мембран. В липидном бислое гидрофобные цепочки жирных кислот ориентированы практически параллельно друг другу, в результате чего образуется достаточно жесткая структура. При повышении температуры гидрофобный слой переходит из упорядоченного состояния в неупорядоченное, и образуется более жидкая, текучая система. Температура, при которой вся структура претерпевает переход из упорядоченного состояния в беспорядочное, называется температурой перехода. Более длинные и более насыщенные жирнокислотные цепи обладают более высокой температурой перехода, т.е. для повышения текучести образованной ими структуры необходима более высокая температура. Наличие ненасыщенных связей в цис-конфигурации приводит к повышению текучести бислоя из-за снижения компактности упаковки цепей без изменения гидрофобности (рис. 42.3). Фосфолипиды клеточных мембран обычно содержат по крайней мере одну ненасыщенную жирную кислоту, имеющую по крайней мере одну двойную связь в цис-положении.
Холестерол играет роль молекулярного модификатора мембран, включение которого приводит к образованию состояний с промежуточной текучестью. Если ацильные боковые цепи находятся в неупорядоченном состоянии, то холестерол вызывает их конденсацию; если же они образуют какую-то кристаллоподобную структуру, то холестерол переводит ее в неупорядоченное состояние. При высоком отношении холестерол/липид фазовый переход вообще не происходит.
Текучесть мембраны сильно влияет на ее функционирование. При увеличении текучести мембрана становится более проницаемой для воды и других малых гидрофильных молекул, растет скорость латеральной диффузии интегральных белков. Если активный центр интегрального белка, осуществляющий некую функцию, располагается исключительно в гидрофильной его части, то изменение текучести липидов, вероятно, не скажется слишком сильно на активности белка. Но если белок выполняет транспортную функцию и транспортный компонент пересекает мембрану, то изменения свойств липидной фазы могут привести к значительному изменению скорости транспорта. Превосходным примером является зависимость функционирования инсулинового рецептора от текучести мембран (гл. 51). Когда концентрация ненасыщенных жирных кислот в мембране растет (при культивировании клеток в среде, богатой этими соединениями), увеличивается текучесть, а это приводит к тому, что рецептор связывает больше инсулина.
Текучесть мембраны и соответственно латеральная подвижность могут быть неодинаковыми в разных ее участках. Например, в плоскости мембраны могут возникать белок-белковые взаимодействия, приводящие к образованию жесткого белкового матрикса в отличие от обычного липидного матрикса. Такие области белкового матрикса могут сосуществовать с обычным липидным матриксом в одних и тех же мембранах. Примерами такого тесного соседства различных матриксов являются области щелевых контактов, плотных контактов, а также бактериородопсинсодержащие фрагменты пурпурных мембран галобактерий.
Некоторые латеральные белок-белковые взаимодействия опосредуются периферическими белками; например, образуются сшивки через антитела и лектины и формируются так называемые кэп-структуры на поверхности мембраны. Таким образом, периферические белки, участвуя в специфических взаимодействиях, могут ограничивать подвижность интегральных белков внутри мембраны.