Биохимия человека Том 2 - Марри Р. 1993
Биохимия внутри- и межклеточных коммуникаций
Действие гормонов
Механизм действия гормонов I группы
Общая схема действия гормонов этой группы показана на рис. 44.1. Их липофильные молекулы диффундируют сквозь плазматическую мембрану любых клеток, но только в клетках-мишенях они находят свой специфический рецептор, имеющий высокую степень сродства к гормону. Образуется комплекс гормон — рецептор, который далее подвергается «активации». В результате этой реакции, зависящей от температуры и присутствия солей, меняется величина, конформация и поверхностный заряд комплекса, и он приобретает способность связываться с хроматином. Вопрос о том, где происходит образование и «активация» комплекса — в цитоплазме или ядре, — остается спорным, но он не очень существен для понимания процесса в целом. Гормонрецепторный комплекс связывается со специфической областью ДНК и активирует либо инактивирует специфические гены. В результате избирательного воздействия на транскрипцию генов и синтез соответствующих мРНК происходит изменение содержания определенных белков, что сказывается на активности тех или иных процессов метаболизма. Эффект каждого из гормонов описываемой группы совершенно специфичен; как правило, их влияние сказывается менее чем на 1% белков или мРНК клетки-мишени. Здесь мы обсуждаем ядерный механизм действия стероидных и тиреоидных гормонов, поскольку этот механизм хорошо изучен. Однако имеются данные о прямом эффекте указанных гормонов на компоненты цитоплазмы и различные органеллы.
Было показано воздействие эстрогенов и глюкокортикоидов на деградацию мРНК; известно также, что глюкокортикоиды оказывают влияние на посттрансляционный процессинг некоторых белков. Но все же большая часть данных указывает на то, что основной эффект этих гормонов проявляется на уровне транскрипции генов. Хотя биохимический механизм транскрипции генов в клетках млекопитающих не вполне ясен, тем не менее можно представить себе в общей модели те структурные компоненты, которые необходимы для проявления регулирующего эффекта стероидных и тиреоидных гормонов на этот процесс (рис. 44.2). Транскрибируемые гены должны находиться в участках «открытого», т. е. транскрипционно-активного, хроматина (изображено в виде вздутия на рис. 44.1), о чем свидетельствует их чувствительность к гидролизу ДНКазой I. Такие гены, судя по полученным к настоящему времени данным, содержат по крайней мере два разных регуляторных элемента (сайты регуляции), расположенных в последовательности ДНК, примыкающей к 5'-концу сайта инициации транскрипции (рис. 44.2). Первый из них — премоторный элемент (ПЭ) — универсален, поскольку в той или иной форме имеется во всех генах. Он определяет место прикрепления РНК-полимеразы II к ДНК, а следовательно, и точность начала транскрипции (начала считывания ДНК) (см. гл. 41).
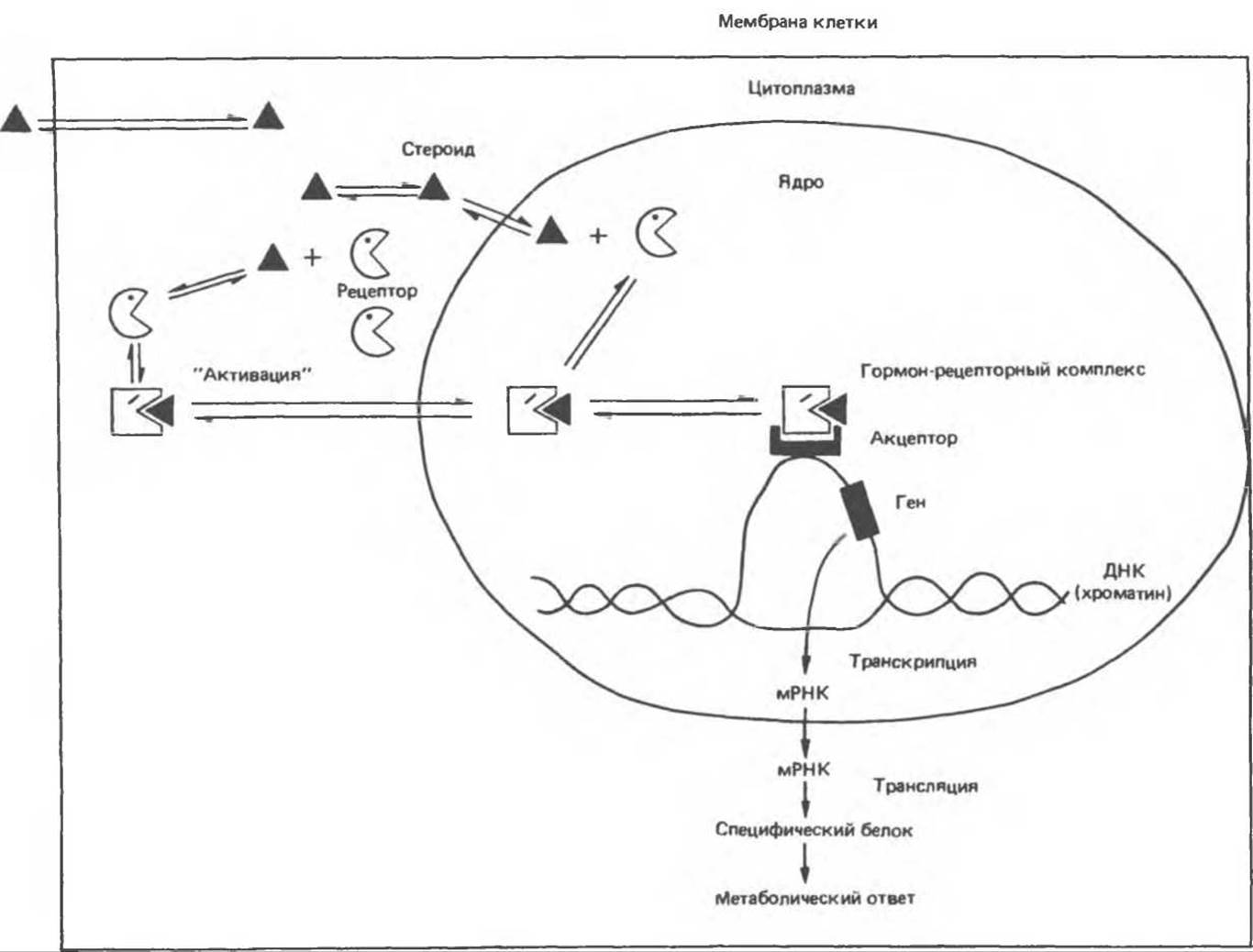
Рис 44.1. Стероидный гормон связывается с внутриклеточным рецептором и вызывает изменение его конформации. Далее этот комплекс связывается со специфической областью на хроматине, что приводит к активации ограниченного числа генов. Тиреоидные гормоны связываются с рецептором, составляющим часть хроматинового комплекса. В остальном механизм действия этих гормонов, по-видимому, одинаков.
Второй элемент — гормон-чувствительный (ГЧЭ) — выявлен во многих генах, регулируемых стероидными гормонами. Он локализован несколько дальше от 5'-конца, чем ПЭ, и может состоять из нескольких отдельных компонентов. Считается, что ГЧЭ модулирует частоту инициации транскрипции и в меньшей степени зависит от положения и ориентации (по сравнению с ПЭ). В этом отношении он похож на энхансерные элементы, обнаруженные в других генах (см. гл. 41). Как правило, ГЧЭ выявляется на несколько сотен нуклеотидов выше сайта инициации транскрипции, но точная локализация варьирует от гена к гену. В некоторых случаях этот элемент расположен в самом транскрибируемом гене.

Рис. 44.2. Структурные компоненты, участвующие в стероидной регуляции транскрипции генов.
Для идентификации ГЧЭ необходимо, чтобы он связывал гормон-рецепторный комплекс с большим сродством, чем остальная ДНК в ядре или ДНК из другого источника. Такое специфическое связывание было действительно продемонстрировано. Кроме того, ГЧЭ должен передавать дальше свой ответ на гормон. Чтобы это проверить, предполагаемую регуляторную последовательность ДНК «сшивают» с маркерным геном. Обычно в такие составные гены включают те маркеры, которые в нормальных условиях не подвержены влиянию гормона. В качестве маркерных генов используют чаще всего гены глобина, тимидинкиназы или бактериальной хлорамфеникол-ацетилтрансферазы. Образовавшиеся при слиянии составные гены переносят в клетку-мишень, и если после этого обнаруживается, что гормон начинает регулировать транскрипцию маркера, то наличие функционально активного ГЧЭ можно считать доказанным. Использование этой техники позволяет точно определить положение ГЧЭ, его ориентацию и эффект замещения оснований. Конкретный механизм того, как влияет на транскрипцию взаимодействие гормон-рецепторного комплекса с ГЧЭ, исследуется очень интенсивно. Предположительно регуляция осуществляется на уровне инициации транскрипции, но возможен эффект и на процессы элонгации и терминации. Высказывались предположения, что регуляторные сайты локализованы в самом гене или вне его в положении выше 5' от сайта инициации или ниже 3'. Наконец, возможно также участие и транс-активных регуляторных механизмов (т. е. воздействие со стороны другой хромосомы).