Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Биохимическая генетика и синтез нуклеиновых кислот и белков
Репликация ДНК
Репликация вирусной ДНК
Пытаясь найти по возможности более простые системы для изучения синтеза ДНК, многие исследователи обратились к мелким ДНК- содержащим вирусам типа —Х174 и М13. Они не обошли при этом вниманием бактериофаги, снабженные отростками: фаги λ, Т7 и Т4, а также плазмиду колицина Е-1. Преимущество этих систем состоит в том, что для них легче смоделировать репликацию ДНК в клеточных экстрактах, а кроме того, ДНК вирусов и плазмид хорошо изучены с генетической точки зрения. Во многих случаях репликация зависит как от генов вируса, так и от генов клетки-хозяина. Так, например, мутации генов dnaB, D, Е, F и G приводят к потере способности поддерживать рост фага λ точно так же, как и в случае, когда инактивированы ts-гены. Вместе с тем фаг λ сохраняет способность к репликации в бактериях с мутантными генами A и С. Многие вирусы, в том числе Т-четные фаги, содержат гены, кодирующие синтез своих собственных специфических ДНК-полимераз и других белков, необходимых для репликации.
а. Репликативные формы
Было установлено, что на первом этапе репликации вирусов — X и М13 присходит превращение одноцепочечной замкнутой кольцевой ДНК инфицирующей вирусной частицы в кольцевую двухцепочечную реплинативную форму (RF). Двухцепочечные кольцевые молекулы далее несколько раз подвергаются репликации, в результате чего образуется большое число кольцевых молекул RF, играющих роль матриц для синтеза большого числа одноцепочечных вирусных (+)-ДНК, из которых образуются зрелые вирусы. Оба этапа — превращение одноцепочечной ДНК в репликативную форму и удвоение последней — в настоящее время интенсивно исследуются [201]. Для осуществления первого этапа у вирусов —Х и М13 геном бактерии-хозяина должен обязательно содержать активный ген ДНК-полимеразы III (dnaE)1) Тем не менее оказалось, что если при проведении реакции в пробирке ввести в нее предварительно очищенную ДНК-полимеразу III, то она не проявляет активности. Дальнейшие попытки получить чистый фермент привели к выделению нового продукта гена dnaE, который получил название ДНК-полимеразы III*. Этот фермент является, судя по всему, более высокомолекулярным полимером, чем ДНК-полимераза III [202]. Для осуществления репликации необходим, однако, еще один белок с мол. весом 77000, известный под названием кополимеразы III*. Наиболее активен тетрамер кополимеразы III* и полимеразы III [202]. Для функционирования системы необходимо также наличие спермидина и АТР (во время инициации процесса полимеризации АТР расщепляется до ADP и Pi), а также всех четырех дезоксирибонуклеозидтрифосфатов.
б. РНК-затравки
Исследователей репликации вирусов М13 и —Х ожидал еще один сюрприз. Оказалось, что для образования RF-ДНК необходима РНК-полимераза. Это послужило одним из многих доводов в пользу предположения, согласно которому для того, чтобы начать синтез ДНК необходим небольшой фрагмент (затравка) РНК [уравнение (15-3)]. Аналогичные наблюдения были сделаны при изучении репликации ДНК плазмиды колицин Е-1. Этот процесс чувствителен к рифампицину — специфическому ингибитору РНК-полимеразы (дополнение 15-А). Для репликации ДНК наряду с дезоксирибонуклеозидтрифосфатами необходимы также четыре рибонуклеозидцрифосфата [203].
В результате последующих исследований было установлено, что в кольцевой ДНК плазмид присутствуют фрагменты РНК, которые могут содержаться также и в ДНК вирусов и Е. coli. В случае фага —Х синтез этой РНК обеспечивает специальная резистентная к рифампицину РНК-полимераза с мол. весом приблизительно 64 000, синтез которой закодирован в гене dnaG Е. coli. Возможно, что именно этот фермент необходим для нормального первичного синтеза ДНК у Е. coli [204]. В удалении РНК-затравки определенную роль может играть специальная рибонуклеаза РНКаза Н, специфически расщепляющая цепь PHK из комплекса РНК—ДНК. В результате действия РНКазы Н в цепи будет образовываться промежуток, ликвидировать который может ДНК-полимераза [205]. Однако вопрос о том, какой фермент на самом деле удаляет РНК-затравку, остается открытым. В опытах in vitro РНКаза Н удаляет РНК не полностью. Не исключено, что РНК- затравка удаляется за счет 5'-экзонуклеазной активности ДНК-полимеразы I (рис. 15-30) [205а].
Репликация фага — X требует также продуктов генов dna В, С, D и G, «расплетающего белка» (разд. Д,5,в) и двух других «факторов».
в. «Расплетающие» белки
При генетическом анализе процесса репликации ДНК фага Т4 оказалось, что для образования вирусной ДНК в клетках Е. coli необходимо по меньшей мере пять вирусных генов. Один из них, а именно ген 43, кодирует синтез ДНК-полимеразы фага ТА, тогда как ген 32 кодирует белок, известный под названием ДНК-расплетающего белка [206]. Этот белок, обладая большим сродством к одноцепочечной ДНК, чем к двухцепочечной, связывается с концевым одноцепочечным участком двойной спирали ДНК, расплетая ее и делая доступными пуриновые и пиримидиновые основания матричной цепи. В результате генетических исследований было установлено, что этот белок необходим как для репликации, так и для генетической рекомбинации. Был выделен аналогичный белок, синтез которого индуцируется фагом Т7; другой белок этого же типа удалось выделить из неинфицированной Е. coli [207, 208]. У эукариот также имеются белки, связывающиеся с ДНК [208а]. Истинная функция этих «расплетающих» (белков на молекулярном уровне все еще не установлена. Таким образом, дальнейшие исследования процесса репликации ДНК должны быть направлены на идентификацию необходимых белков и на изыскание способов осуществления репликации в пробирке при помощи систем, обеспечивающих реализацию этого процесса в живой клетке.
г. Репликация вирусных двухцепочечных RF-молекул
Известно, что для инициации процесса репликации ДНК фага —Х необходимо наличие в геноме фага специфического гена А. Недавно было показано, что этот ген детерминирует синтез белка с мол. весом 56 000 — специфической эндонуклеазы надрезающей вирусную цепь RF-формы, что необходимо для начала процесса репликации [209]. По-видимому, после появления такого разрыва стимулируется синтез небольшого участка РНК-затравки. Репликация ДНК протекает в большинстве случаев в двух направлениях (разд. Д, 2), однако репликативная форма —X образуется, вероятно, только в одном направлении по механизму «разматывающегося рулона» (rolling circle) [210]. В соответствии с этим механизмом [уравнение (15-9)], по мере того как вновь образующаяся цепь вирусной ДНК синтезируется вдоль комплементарной (минус) цепи-матрицы, исходная вирусная ДНК (плюс-цепь) вытеснется в виде одноцепочечного «хвоста».
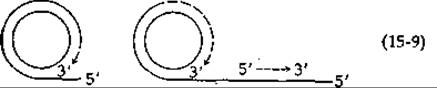
После этого образуется цепь (возможно, в форме отдельных фрагментов), комплементарная одноцепочечному «хвосту». При полном обороте кольца получается цепь вирусной ДНК в два раза длиннее обычной. Процесс репликации завершается ее расщеплением при помощи соответствующей эндонуклеазы и сшиванием комплементарных нитей в кольцо лигазой.