БИОХИМИЯ - Л. Страйер - 1984
ТОМ 1
ЧАСТЬ I. КОНФОРМАЦИЯ И ДИНАМИКА
ГЛАВА 7. МЕХАНИЗМ ДЕЙСТВИЯ ФЕРМЕНТОВ: ЛИЗОЦИМ И КАРБОКСИПЕПТИДАЗА
7.2. Трехмерная структура лизоцима
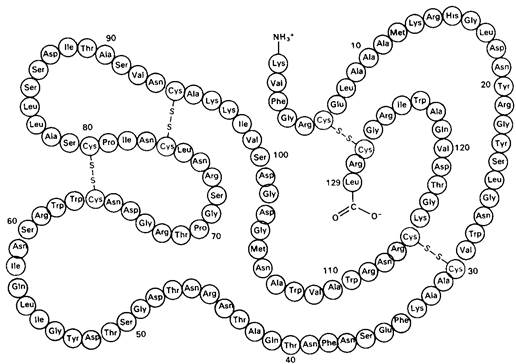
Лизоцим-относительно небольшой фермент. Лизоцим, выделенный из белка куриных яиц, где он содержится в большом количестве, представляет собой одну полипептидную цепь из 129 аминокислотных остатков и массой 14,6 к Да. В молекуле лизоцима имеется четыре поперечных дисульфидных мостика, обусловливающих стабильность фермента. Последовательность аминокислот в лизоциме показана на рис. 7.5.
Рис. 7.5. Последовательность аминокислот в лизоциме из белка куриных яиц. Красным показаны остатки, входящие в состав активного центра
В 1965 г. Дэвид Филлипс (D. Phillips) с сотрудниками определили трехмерную структуру лизоцима. Так впервые была получена карта высокого разрешения для белка, обладающею ферментативной активностью. Молекула лизоцима оказалась компактной, приближенно эллипсоидной формы размерами 45 х 30 х 30 А. Как показано на рис. 7.7, укладка полипептидной цепи носит очень сложный характер. В ней содержится гораздо меньше α-спиралей, чем в миоглобине и гемоглобине. В некоторых участках полипептидная цепь имеет вытянутую конформацию. В одном из этих участков цепь делает поворот и идет в обратном направлении параллельно самой себе; при этом два параллельных тяжа соединяются водородными связями, образующимися между пептидными группами. Такие участки «шпильки для волос» подобны упоминавшимся выше антипараллельным β-складчатым слоям -регулярно повторяющимся участкам вторичной структуры в белке шелка. Внутренняя часть молекулы лизоцима, подобно молекулам миоглобина и гемоглобина, почти полностью неполярна. Очевидно, что, как и в случае большинства других белков, гидрофобные взаимодействия играют важную роль в образовании третичной структуры лизоцима.
7.3. Поиски активного центра лизоцима
Детальное воспроизведение трехмерной структуры лизоцима еще не раскрывало механизма его каталитического действия. Более того, рассматривая карту электронной плотности, трудно было определить хотя бы локализацию активного центра. В отличие от миоглобина и гемоглобина в лизоциме нет простетической группы, т.е. нет встроенного маркера активного центра. В итоге основную информацию, необходимую для идентификации активного центра, определения способа связывания субстрата и выяснения механизма ферментативного действия, удалось получить только при использовании ингибиторов, а именно при рентгеноструктурном анализе комплексов лизоцима с ингибиторами. Если трехмерная структура белка известна, то определить методом рентгеноструктурного анализа способ связывания с ним небольших молекул обычно уже нетрудно. Эти опыты легко выполнимы, потому что кристаллы белка очень пористы. Даже относительно большие молекулы ингибитора способны диффундировать по каналам между молекулами белка, попадая в участки специфического связывания. Изменение электронной плотности, обусловленное встраиванием дополнительной молекулы, можно рассчитать непосредственно по интенсивности рефлексов (используя данные, полученные предварительно на нативном белке) при условии, что структура кристалла не претерпела существенных изменений. Такой подход получил название разностного метода Фурье.
Рис. 7.6. Рентгенограмма кристалла лизоцима

В идеале следовало бы использовать разностный метод Фурье для определения структуры фермент-субстратного (ЕS) комплекса в процессе катализа. Однако в обычных условиях превращение связанного субстрата в продукт реакции происходит значительно быстрее, чем диффузия новых молекул субстрата в кристалл. В некоторых случаях это осложнение удается устранить замедлением каталитического процесса, что достигается путем сильного охлаждения кристалла (например, до — 50 С). Этот экспериментальный подход получил название криоэнзимологии. Существует и другой подход: исследуют комплекс фермента с таким аналогом субстрата, который либо совсем не подвергается никаким превращениям, либо эти превращения происходят очень медленно. Для лизоцима таким аналогом субстрата оказался тример N-ацетилглюкозамина (три-NAG, или NAGз), структура которого показана на рис. 7.8. Олигомеры N-ацетилглюкозамина, содержащие менее 5 остатков, гидролизуются крайне медленно или не гидролизуются совсем. Тем не менее они связываются с активным центром фермента. Именно поэтому три-NАG является мощным конкурентным ингибитором лизоцима.
Рис. 7.7. Трехмерная структура лизоцима. Показаны только α-углеродные атомы
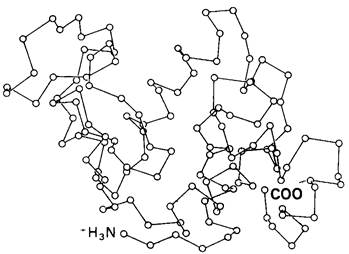
Рис. 7.8. Формула три-N-ацетилглюкозамина (три-NАG, или NАG3) - конкурентного ингибитора лизоцима