БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 12. ГЛИКОЛИЗ
12.7. Образование пирувата и генерирование второй молекулы АТР
Мы подошли теперь к последней стадии гликолиза. Она осуществляется в три этапа, в результате которых происходит превращение 3-фосфоглицерата в пируват и образование второй молекулы АТР.
Первая из этих реакций представляет собою внутримолекулярную перестройку. При превращении 3-фосфоглицерата в 2-фосфоглицерат, катализируемом фосфо- глицеромутазой, происходит перемещение фосфорильной группы. Вообще мутазой называют фермент, катализирующий внутримолекулярные перемещения химических групп, таких, как фосфорильная группа.
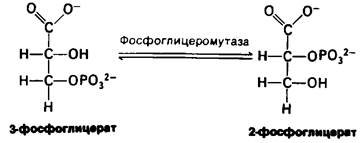
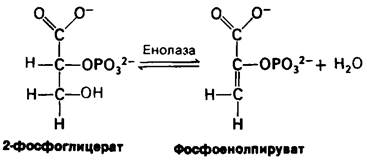
Вторая реакция состоит в образовании енола в результате дегидратации 2-фосфо- глицерата. Образование фосфоенолпирувата катализируется енолазой. Потенциал переноса фосфорильной группы в результате реакции дегидратации значительно повышается. Енолфосфат характеризуется высоким потенциалом переноса фосфорильной группы, тогда как в случае фосфатного эфира обычного спирта этот потенциал находится на низком уровне. Причины указанного различия мы рассмотрим позднее.
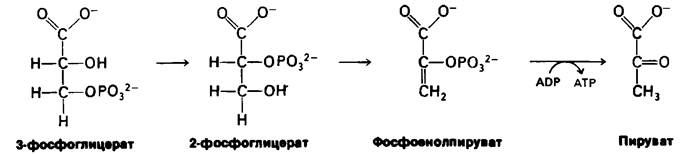
Последняя реакция -образование пирувата с одновременным образованием АТР. Перенос фосфорильной группы от фосфоенолпирувата к ADP катализируется пируваткиназой. Это фосфорилирование не является окислительным в противоположность реакции, катализируемой глицеральдегид-3-фосфат-дегидрогеназой.
12.8. Выход энергии при превращении глюкозы в пируват
Суммарная реакция превращения глюкозы в пируват имеет следующий вид:
Глюкоза + 2 Pi + 2ADP + 2NAD+ → 2 Пируват + 2АТР + 2NADH + 2Н+ + 2Н2O.
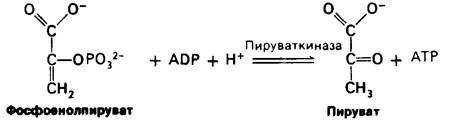
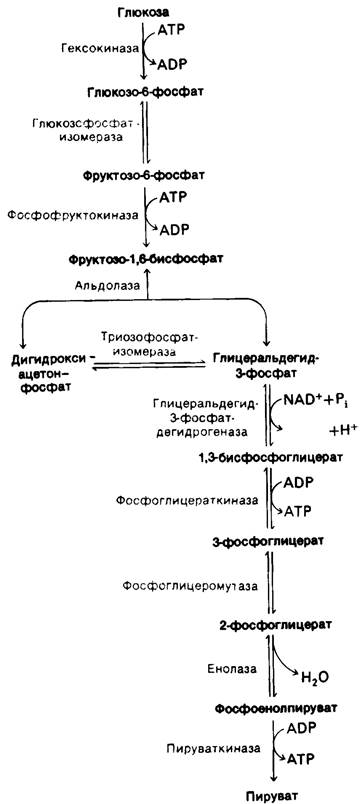
Таким образом, в ходе превращения глюкозы в пируват происходит образование двух молекул АТР. Суммарный перечень этапов, на которых происходит потребление или образование АТР, приведен в табл. 12.1. Напомним, что из фруктозо-1,6-бисфосфата образуются два трехуглеродных компонента. Реакции гликолиза суммированы в табл. 12.2 и на рис. 12.8.
Таблица 12.1. Потребление и образование АТР при гликолизе

Таблица 12.2. Реакции гликолиза

Рис. 12.8. Гликолитический путь
12.9. Фосфофруктокиназа - ключевой фермент в регуляции гликолиза
Гликолитический путь играет двоякую роль: он приводит к генерированию АТР в результате расщепления глюкозы, и он же поставляет строительные блоки для реакций синтеза, таких, как образование жирных кислот с длинной цепью. Скорость превращения глюкозы в пируват регулируется таким образом, чтобы удовлетворять этим двум главным потребностям клетки. В метаболических путях ферменты, катализирующие по существу необратимые реакции, играют роль потенциальных участков контроля. При гликолизе реакции, катализируемые гексокиназой, фосфофруктокиназой и пируваткиназой, фактически необратимы и, по-видимому, могут играть и регуляторную, и каталитическую роль. Все три фермента действительно являются теми участками регуляции метаболического пути, на котором осуществляется регуляция гликолитического процесса.
Фосфофруктокиназа - наиболее важный регуляторный компонент гликолиза. Этот тетрамерный фермент ингибируется высокими концентрациями АТР, снижающими его сродство к фруктозо-6-фосфату. Если реакция протекает в присутствии высоких концентраций АТР, то кинетика фосфофрукто- киназной реакции описывается не гиперболической кривой, а сигмоидной (рис. 12.9). Этот аллостерический эффект усиливается при связывании АТР с высокоспецифичным регуляторным центром, локализованным отдельно от каталитического центра. Ингибиторный эффект АТР обращается под действием АМР. Следовательно, активность фермента возрастет при снижении отношения [АТР]/[АМР]. Другими словами, гликолиз стимулируется в условиях низкого энергетического заряда клетки. Как уже упоминалось, гликолиз поставляет также углеродный скелет для процессов биосинтеза. Следовательно, регуляторное действие на фосфофруктокиназу будут, очевидно, оказывать также сигналы об избытке или недостатке строительных блоков. Действительно, фосфофруктокиназа ингибируется цитратом, ранним промежуточным продуктом в цикле трикарбоновых кислот (разд. 13.2). Высокое содержание цитрата означает, что соединения, играющие при биосинтезе роль предшественников, присутствуют в избытке и дополнительного распада глюкозы для этой цели не требуется. Цитрат подавляет активность фосфофруктокиназы, усиливая ингибиторное действие АТР. Таким образом, фосфофруктокиназанаиболее активна, когда клетка нуждается и в энергии. и в строительных блоках. о чем сигнализируют низкое значение отношения [АТР]/[АМР] и низкое содержание цитрата. Фермент обладает умеренной активностью в условиях, когда существует потребность либо в энергии, либо в углеродном скелете. Если оба пни фактора присутствуют в избытке, активность фосфофруктокиназы падает почти до нуля.
Рис. 12.9. Аллостерическая регуляция фосфофруктокиназы. Высокая концентрация АТР ингибирует фермент, понижая его сродство к фруктозо-6-фосфату. АМР снижает, а цитрат усиливает ингибирующее действие АТР

Гексокиназа и пируват киназа также участвуют в регуляции скорости гликолиза. Пируваткиназа мышц и печени аллостерически ингибируется АТР, и. таким образом, превращение фосфоенолпиру вата в пируват блокируется в условиях высокою энергетического заряда. Гексокиназа аллостерически ингибируется глюкозо-6-фосфатом. Содержание фруктозо-6-фосфата повышаема при подавлении активности фосфофруктокиназы; при этом происходит соответствующее увеличение количества глюкозо-6-фосфата, который находится в равновесии с фруктозо-6-фосфатом. Следовательно, ингибирование фосфофруктокиназы высоким отношением |АТР]/[АМР] или высоким содержанием цитрата приводит к ингибированию гексокиназы. В печени глюкоза фосфорилируется в глюкозо-6-фосфат даже при высокой концентрации последнего, что обусловливается присутствием глюкокиназы - фермента, отличного от гексокиназы. Глюкокиназа характеризуется высоким значением КМ, для глюкозы, и, следовательно, она активна только в условиях избытка глюкозы. Роль глюкокиназы состоит в том, чтобы поставлять глюкозо-6- фосфат для синтеза гликогена, резервной формы глюкозы (гл. 16). Высокая Км глюкокиназы печени дает возможность мозгу и мышцам первыми использовать глюкозу при ее ограниченном поступлении.
Почему ведущую роль в гликолизе играет фосфофруктокиназа, а не гексокиназа? Это станет понятным, если вспомнить, что глюкозо-6-фосфат является промежуточным продуктом не только гликолитического пути. Он может также превращаться в гликоген или окисляться по пентозофосфатному пути (гл. 15) с образованием NADPH. Первая необратимая реакция, присущая только гликолитическому пути, названная решающим этапом, зто фосфорилирование фруктозо-6-фосфата во фруктозо-1,6-бисфосфат. Отсюда вытекает значение фосфофруктокиназы как основного фактора регуляции гликолиза. Вообще фермент, катализирующий решающий этан в метаболической последовательности, всегда является наиболее важным регуляторным элементом данного пути.
Рис. 12.10. Электронная микрофотография дрожжевой клетки