БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 14. ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ
14.12. Полное окисление глюкозы дает 36 АТР
Теперь мы можем рассчитать количество АТР, образующееся при полном окислении глюкозы (табл. 14.4). Суммарная реакция:
Глюкоза + 36ADP + 36Pi + 36Н+ + 6O2 → 6СO2 + 36АТР + 42Н2O.
Таблица 14.4. Выход АТР при полном окислении глюкозы

Отношение Р:0 равно 3, поскольку образуется 36 молекул АТР. а потребляется 12 атомов кислорода. Большая часть АТР, 32 молекулы из 36, генерируется путем окислительного фосфорилирования.
Общая эффективность генерирования АТР достигает высокого уровня. При окислении глюкозы при стандартных условиях высвобождается 686 ккал:
Глюкоза + 6O2 → 6СO2 + 6Н2O
∆G0' = - 686 ккал.
Свободная энергия, запасенная в 36 АТР, составляет 263 ккал, поскольку ∆G0' для гидролиза АТР равно — 7,3 ккал. Отсюда термодинамическая эффективность образования АТР из глюкозы достигает при стандартных условиях 263/686. или 38°0.
Дыхательный коэффициент (RQ). часто используемый показатель при исследованиях метаболизма на целом организме, определяется следующим путем:
![]()
Для полного окисления углеводов RQ равен 1. Для жиров и белков он составляет соответственно 0.71 и 0.80. Таким образом. RQ может быть использован как показатель относительного использования организмом углеводов, жиров и белков.
4.13. Скорость окислительного фосфорилирования определяется по потребности в АТР
При большинстве физиологических состояний перенос электронов тесно сопряжен с фосфорилированием. Электроны обычно переносятся по электронтранспортной цепи лишь при условии одновременного фосфорилирования ADP в АТР. Окислительное фосфорилирование требует поступления NADH (или другого источника электронов с высоким потенциалом), О2, ADP и Pi. Наиболее важным фактором в определении скорости окислительного фосфорилирования является содержание ADP. При добавлении ADP скорость поглощения кислорода тканевым гомогенатом значительно возрастает и затем, когда весь добавленный ADP превратится в АТР, возвращается к исходному уровню (рис. 14.12).
Рис. 14.12. Дыхательный контроль. Электроны переносятся на О2 только при условии фосфорилирования ADP в АТР
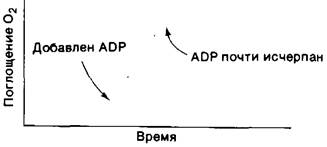
Регуляция скорости окислительного фосфорилирования содержанием ADP называется дыхательным контролем. Физиологическое значение этого регуляторного механизма очевидно. Содержание ADP возрастает при потреблении АТР, и, таким образом, окислительное фосфорилирование оказывается сопряженным с использованием АТР. При отсутствии потребности в синтезе АТР переноса электронов от топливных молекул на О2 не происходит.
14.14. Динитрофенол разобщает окислительное фосфорилирование путем разрушения протонного градиента
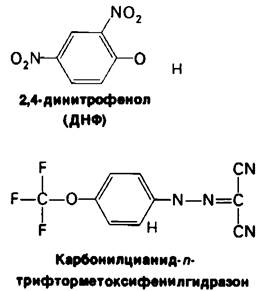
Тесное сопряжение между переносом электронов и фосфорилированием нарушается под действием 2,4-динитрофенола (ДНФ) и некоторых других кислотных ароматических соединений (рис. 14.13). Эти соединения переносят протоны через внутреннюю митохондриальную мембрану. Перенос электронов от NADH к О2 в присутствии таких разобщителей протекает нормально, но образования АТР митохондриальной АТРазой не происходит, поскольку исчезает протонодвижущая сила, обусловливающая перенос протонов через внутреннюю мембрану митохондрий. Потеря дыхательного контроля приводит к тому, что поглощение О2 и окисление NАDН повышается. В то же время ДНФ не оказывает влияния на субстратное фосфорилирование. В силу своего специфического действия на дыхательную цепь ДНФ и другие разобщители служат ценным инструментом при исследованиях обмена веществ.
Рис. 14.13. Формулы двух разобщителей окислительного фосфорилирования. Эти растворимые в липидах соединения могут переносить протоны через внутреннюю митохондриальную мембрану. Отщепляющийся протон показан красным цветом
Разобщение окислительного фосфорилирования может быть биологически полезным. Оно представляет собою способ генерирования тепла для поддержания температуры тела у зимнеспящих животных, у некоторых новорожденных животных и у млекопитающих, адаптированных к холоду. Для этого процесса термогенеза специализирована бурая жировая ткань, очень богатая митохондриями. В качестве разобщителей в ней выступают жирные кислоты, высвобождение которых в свою очередь регулируется норадреналином. Таким образом, степень разобщения окислительного фосфорилирования в бурой жировой ткани находится под гормональным контролем. Митохондрии в этой ткани могут выполнять функцию генераторов АТР, или миниатюрных обогревательных печей.
Существует интересное сообщение о 38-летней женщине, которая была не в состоянии выполнять продолжительную физическую работу. Основной обмен у нее более чем вдвое превышал нормальный уровень, но функция щитовидной железы не была нарушена. Биопсия мышцы показала, что митохондрии в ней очень многообразны и атипичны по структуре. При биохимических исследованиях выяснилось, что эти митохондрии нс подвержены дыхательному контролю. NADH в них окислялся независимо от присутствия ADP. Другими словами, в них отсутствовало тесное сопряжение окисления и фосфорилирования. Отношение Р : O было ниже нормы. Таким образом, у этой больной значительная часть энергии топливных молекул превращалась не в АТР, а в тепло. Природа молекулярного дефекта в таких митохондриях пока не расшифрована.