БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 18. РАЗРУШЕНИЕ АМИНОКИСЛОТ И ЦИКЛ МОЧЕВИНЫ
18.6. Наследственные ферментные нарушения цикла мочевины приводят к гипераммонемии
Высокая концентрация Nтоксична для человека. Главный путь удаления NН4+ - это синтез мочевины в печени. Полное блокирование какой-либо из стадий цикла мочевины в печени, по-видимому, несовместимо с жизнью, потому что другого известного пути синтеза мочевины не существует. Диагностированы наследственные нарушения, вызываемые частичным блокированием одной из реакций цикла мочевины. Общим признаком таких нарушений является повышенное содержание в крови (гиперам- монемия). Почти полная недостаточность какого-либо из ферментов цикла мочевины вызывает кому и приводит к смерти вскоре после рождения. Частичная недостаточность этих ферментов вызывает задержку умственного развития, летаргию, временами рвоту. Малобелковая диета приводит к снижению содержания аммиака в крови и к улучшению клинической картины при мягких формах этих наследственных нарушений.
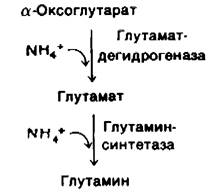
Почему высокое содержание N4+ токсично? Возможно, это объясняется тем, что высокая концентрация иона аммония сдвигает равновесие реакции, катализируемой глутамат-дегидрогеназой, в сторону образования глутамата, а это приводит к истощению α-оксоглутарата. Реакция может быть далее. стимулирована путем включения NH4+ в глутамат с образованием глутамина (разд. 21.2). Истощение а-оксоглутарата, промежуточного продукта цикла трикарбоновых кислот, приводит к уменьшению скорости образования АТР. Мозг отличается высокой чувствительностью к снижению содержания АТР.
18.7. Судьба атомов углерода распавшихся аминокислот
Мы рассмотрели ряд реакций, приводящих к удалению из аминокислот α-аминогруппы и превращению ее в мочевину. Теперь мы обратимся к судьбе оставшихся углеродных скелетов. Стратегия разрушения аминокислот состоит в образовании главных промежуточных продуктов обмена веществ, которые могут превращаться в глюкозу или окисляться в цикле трикарбоновых кислот. В самом деле, углеродные скелеты разнообразного набора из двадцати аминокислот направленно превращаются всего в семь молекул: пируват, ацетил-СоА, ацетоацетил-СоА, а-оксоглутарат, сукцинил-СоА, фу Марат и оксалоацетат. Мы сталкиваемся здесь еще с одним примером замечательной экономичности метаболических превращений.
Аминокислоты, распадающиеся с образованием ацетил-СоА или ацетоацетил-СоА, называются кетогенными, поскольку в результате их распада повышается содержание кетоновых тел. Аминокислоты же, распад которых приводит к образованию пирувата, α-оксоглутарата, сукцинил-СоА, фумарата или оксалоацетата, названы глюкогенными. Возможность синтеза глюкозы из этих аминокислот обеспечивается тем обстоятельством, что указанные компоненты цикла трикарбоновых кислот и пируват могут превращаться в фосфоенолпируват и затем-в глюкозу (разд. 15.13.). Напомним, что у млекопитающих отсутствует путь, обеспечивающий непосредственный синтез глюкозы из ацетил-СоА или ацетоацетил- СоА.
Рис. 18.6. Судьба углеродных скелетов аминокислот. Глюкогенные аминокислоты показаны красным цветом, кетогенные- желтым
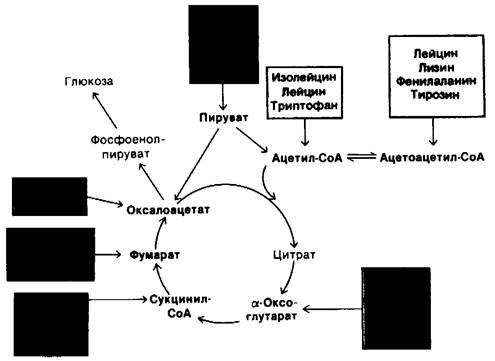
Из основного набора, включающего двадцать аминокислот, только лейцин является исключительно кетогенным. Изолейцин, лизин, фенилаланин, триптофан и тирозин относятся одновременно и к кетогенным, и к глюкогенным аминокислотам. Некоторые из их углеродных атомов появляются в ацетил-СоА или ацетоацетил- СоА, тогда как другие обнаруживаются в потенциальных предшественниках глюкозы. Остальные четырнадцать аминокислот являются чисто глюкогенными.
18.8. Семейство С3-аминокислот; аланин, серин и цистеин превращаются в пируват
Пируват служит «пунктом входа» (в цикл трикарбоновых кислот) для трехуглеродных аминокислот: аланина, серина и цистеина (рис. 18.7). Трансаминирование аланина дает непосредственно пируват:
Аланин + α-Оксоглутарат ⇄ Пируват + Глутамат.
Рис. 18.7. Пируват - «пункт входа» для аланина, серина, цистеина, глицина и треонина
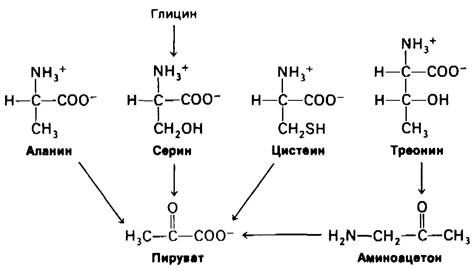
Как указывалось выше (разд. 18.1), глутамат затем подвергается окислительному дезаминированию, приводящему к образованию NН4+ и регенерированию α-оксоглутарата. Суммарная реакция этих превращений описывается уравнением
Аланин + NАD+ → Пируват + NH4+ +NADH+H+
Другая простая реакция в распаде аминокислот представляет собою дезаминирование серина в пируват под действием серин- дегидратазы (разд. 18.3):
Серин → Пируват + NH4+.
Цистеин может превращаться в пируват различными путями с появлением атома серы в Н2S, SО23- или SCN-.
Возможно превращение в пируват и углеродных атомов двух других аминокислот. Глицин может превращаться в серин путем ферментативного присоединения гидроксиметильной группы (разд. 21.5). Из треонина пируват может образовываться через аминоацетон.