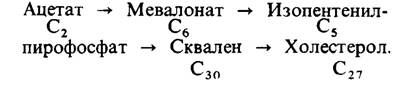БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ III. БИОСИНТЕЗ ПРЕДШЕСТВЕННИКОВ МАКРОМОЛЕКУЛ
ГЛАВА 20. БИОСИНТЕЗ МЕМБРАННЫХ ЛИПИДОВ И СТЕРОИДНЫХ ГОРМОНОВ
20.8. Болезнь Тея-Сакса: наследуемое нарушение расщепления ганглиозидов
Ганглиозиды содержатся в наибольшей концентрации в нервной системе, особенно в сером веществе, где они составляют 6% всех липидов. Ганглиозиды постоянно синтезируются и расщепляются путем последовательного удаления концевых остатков сахара. Катализирующие эти реакции гликозид-гидролазы высокоспецифичны. Расщепление ганглиозидов происходит в лизосомах. Эти органеллы содержат самые разнообразные ферменты расщепления и предназначены для упорядоченного разрушения компонентов клетки.
Нарушения в способности клетки к расщеплению ганглиозидов могут привести к серьезным клиническим последствиям.
Симптомы болезни Тея-Сакса обычно проявляются у ребенка в возрасте до одного года. К характерным ранним симптомам относятся слабость, отставание в развитии, затруднения при кормлении. Через несколько месяцев обычно наступает слепота. Летальный исход при болезни Тея — Сакса обычно наступает до 3 лет. Эта болезнь сопровождается разительными патологическими изменениями в нервной системе: ганглиозные клетки коры головного мозга и некоторых других участков мозга чрезмерно разбухают. Кроме того, на сетчатке видны отчетливые вишнево-красные пятна.
Содержание ганглиозидов в мозгу ребенка, страдающего болезнью Тея-Сакса, сильно повышено. Особенно сильно, во много раз по сравнению с нормой, повышается концентрация ганглиозида GM2. Аномально высокое содержание этого ганглиозида обусловлено недостаточностью фермента, отщепляющего концевой остаток N-ацетилгалактозамина. Фермент, отсутствие (или недостаточная активность) которого обусловливают эту болезнь, специфическая β-N-ацетилгексозаминидаза.
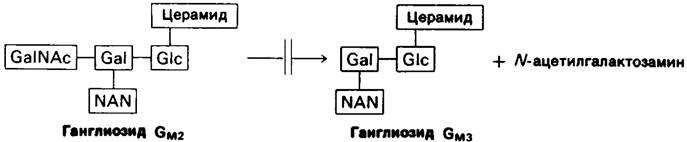
Рис. 20.6. Электронная микрофотография лизосомы
Болезнь Тея-Сакса наследуется как аутосомный рецессивный признак. Частота носителей составляет 1/30 среди американцев еврейского происхождения и 1/300 среди прочих американцев. Следовательно, частота болезни среди американцев еврейского происхождения выше примерно в 100 раз. Болезнь Тея-Сакса можно диагносцировать еще во время эмбрионального развития. Для этого берут пробу амниотической жидкости с помощью амниоцентеза и определяют в ней активность β-N-ацетилгексозаминидазы.
Рис. 20.7. Пространственная модель холестерола
20.9. Холестерол синтезируется из ацетил-кофермента А
Теперь обратимся к синтезу холестерола- стероида, регулирующего текучесть мембран эукариотической клетки (разд. 10.17).

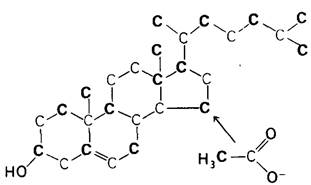
Кроме того, холестерол служит предшественником стероидных гормонов, таких, как прогестерон, тестостерон, эстрадиол и кортизол. Важный шаг в ранних исследованиях синтеза холестерола сделал Конрад Блох (Konrad Bloch) в 40-х годах. Он приготовил ацетат, содержащий меченые атомы углерода, и скормил его крысам. Холестерол, синтезированный в организме этих крыс, содержал изотопную метку, откуда следовало, что ацетат - предшественник холестерола. Действительно, все 27 атомов углерода холестерола происходят из ацетил-СоА. Дальнейший прогресс в изучении синтеза холестерола был достигнут использованием в качестве предшественника ацетата, меченного по метальному или карбоксильному атому углерода. При расщеплении холестерола, синтезированного из ацетата, меченного по одному из атомов углерода, удалось установить происхождение каждого атома в молекуле (рис. 20.8). Это сыграло решающую роль в создании и экспериментальной проверке гипотез о пути синтеза холестерола.
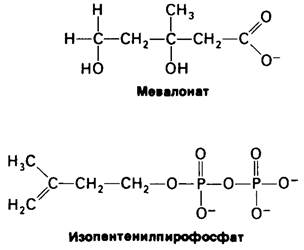
Рис. 20.8. Распределение метки в молекуле холестерола, синтезированного из ацетата, меченного по метильному (показано синим цветом) или карбоксильному (показано красным цветом) атомам углерода
20.10. Мевалонат и сквален-промежуточные продукты синтеза холестерола
Следующим важным достижением было открытие сквалена-С30-углеводорода, промежуточного продукта синтеза холестерола. Сквален состоит из шести изопреновых единиц. Открытие сквалена породило следующую проблему: каким образом синтезируются из ацетата изопреновые единицы.
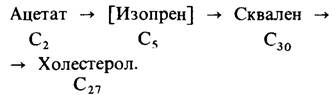
Ответ был неожиданно получен из работ, посвященных исследованию бактериальных мутантов и не имевших никакого отношения к изучению синтеза холестерола. Оказалось, что мевалонат может заменять ацетат, компенсируя его недостаток у мутантов, нуждающихся в ацетате. Открытие мевалоната имело решающее значение для расшифровки пути биосинтеза холестерола, так как вскоре стало ясно, что эта шестиуглеродная кислота может декарбоксилироваться и давать таким образом постулированный пятиуглеродный изопреновый промежуточный продукт. Затем с помощью изотопной метки было показано, что мевалонат действительно служит предшественником сквалена и что он может образовываться из ацетата. Оказалось, что активированный изопреновый промежуточный продукт -изопентенилпирофосфат, который образуется путем декарбоксилирования производного мева- лоната. Итак, схематически синтез холестерола из ацетата можно было представить следующим образом: