Структура и функционирование белков. Применение методов биоинформатики - Джон Ригден 2014
Функциональное разнообразие в элементах упаковки и надсемействах
От способа укладки к функции
Связь между способами укладки и предсказание функций
В этом разделе рассматриваются свойства функций, существование которых можно предположить без использования данных о гомологии, т.е., таких функций, которые возникли в ходе конвергентной эволюции. Вопросы, связанные с изучением функций, установленных на основе информации о гомологии, рассматриваются в разделе 6.3 настоящей главы.
В общем случае, определение структуры и способа укладки белка предоставляет исследователю возможность использовать множество методов предсказания белковых функций на основе структурного анализа, которые были бы недоступны в отсутствие известной белковой структуры. В основе некоторых из этих методов лежит принцип, согласно которому информация о структуре позволяет установить общие гомологии, не очевидные на уровне последовательности (Lee et al. 2007). В других методах структурная информация используется лишь на основе предположения о том, что она является значимой при выполнении белком его молекулярных функций, при этом эволюционный контекст структурных свойств не принимают во внимание. Многие из этих методов описаны в других главах книги (см. главы 7, 8, 10 и 11). В настоящей главе обсуждаются лишь случаи, имеющие непосредственное отношение к информации о способах укладки.
6.2.2.1. Способы укладки с единственной функцией
Структуру белка, определенную недавно, можно использовать для поиска схожих способов укладки среди уже известных белковых структур. При этом используются программы сравнения структур, которые обычно оценивают значимость обнаруженных структурных сходств, используя специфические схемы оценки. Некоторые из этих программ являются общедоступными и в последнее время были протестированы с использованием большого набора данных известных структурных сходств, созданного САТН (Kolodny et al. 2005; Redfem et al. 2007). К числу таких программ принадлежат DALI (Holm and Sander 1996a), FATCAT (Ye and Godzik 2004), SSM (Krissinel and Henrick 2004), CE (Shindyalov and Bourne 1998) и CATHEDRAL (Redfem et al. 2007). В случае, когда новая структура обнаружена в белке с неизвестной функцией, на следующем шаге исследования необходимо оценить, можно ли на основе подобия белковых структур осуществить перенос функциональной аннотации.
Некоторые способы укладки свойственны лишь гомологичным белкам, тогда как другие могут встречаться у белков, эволюционировавших конвергентно, - их принято обозначать гомологичными и аналогичными способами укладки, соответственно (Moult and Melamud 2000). Аналогичным образом, некоторые способы укладки являются однородными с точки зрения функций, тогда как другие характерны для белков, выполняющих весьма разнообразные функции. Принято считать, что гомологичные способы укладки более однородны в функциональном смысле по сравнению с аналогичными (Moult and Melamud 2000). Очевидно, что если способ укладки связан с уникальной функцией X, обнаружение этой укладки в белке с неизвестной функцией прямо приведет к описанию белка с функцией X. Однако на практике ситуация несколько сложнее, поскольку многообразные в функциональном отношении способы укладки можно ошибочно идентифицировать как связанные с единственной функцией из-за специфической выборки.
Так или иначе, известны случаи, когда определение способа укладки способствовало предсказанию функции белка (Moult and Melamud 2000). Например, пространственная структура продукта гена усаС Escherichia coli содержит укладку, которая близка к укладке белков семейства амидогидролаз. Дальнейшие исследования показали, что этот белок имеет каталитический аппарат, близкий к имеющемуся у белков с таким же способом укладки (Colovos et al. 1998; Moult and Melamud 2000). Растет количество случаев успешного предсказания функций белков на основе определения способа укладки с помощью средств структурной геномики, общей целью которой является определение структуры как можно большего числа белков (Adams et al. 2007). В большинстве случаев, однако, успешное предсказание функции белка не является результатом простого определения способа укладки, а связано с сочетанием способа укладки с каким-либо другим свойством, например, мотивом распознавания последовательности или сходством функциональных участков исследуемых белков.
6.2.2.2. Суперучастки
В целом, данные о пространственной структуре весьма полезны при определении функционального участка - подмножества остатков, которые имеют решающее значение для выполнения белком его молекулярной функции. Функциональные участки представлены главным образом участками связывания (совокупность белковых остатков, которые взаимодействуют с лигандами) (Dessailly et al. 2008) или каталитическими участками (совокупность остатков, которые принимают непосредственное участие в ферментативной реакции) (Porter et al. 2004).
Одна из причин, по которым структуры полезны при определении функциональных участков, состоит в том, что последние, как правило, располагаются в наиболее консервативных топологических областях структур. Более того, даже в тех случаях, когда нет убедительных доказательств гомологии белков, имеющих общий способ укладки, функциональные участки располагаются в одних и тех же областях их пространственной структуры. Такие функциональные участки называют суперучастками. Показано, что суперучастки в большом количестве встречаются при аналогичной упаковке (или суперупаковке, см. раздел 6.2.3.1), т.е., при упаковке, которая является общей для негомологичных белков (Russell et al. 1998). На рис. 6.1 представлен хорошо известный пример суперучастка: это каталитический участок белков, для которых характерна упаковка в форме бочонка (ß/a)8. Остатки, принимающие участие в катализе, всегда располагаются на Суконцах ß-тяжей центрального параллельного ß-листа, хотя структура самих ß-тяжей может изменяться (Nagano et al. 2002).
6.2.2.3. Суперукладка
Способ укладки, распространенный среди множества самых разных надсемейств и отличающийся выраженным функциональным разнообразием, называют суперукладкой (Orengo et al. 1994). Элементы суперукладки входят в состав белков, выполняющих большое количество различных функций. Замечательными примерами суперукладки являются способ укладки, напоминающий структуру бочонка TIM (ß/a)8, который встречается у представителей более 25 различных надсемейств (Nagano et al. 2002), а также укладка Россмана, которую можно обнаружить у белков 114 надсемейств САТН (САТН v3.1), многие из которых отличаются функциональным разнообразием. Элементы суперукладки составляют весьма незначительную часть известных способов укладки, однако именно они, по-видимому, являются продуктами большой части известных геномов (Lee et al. 2005). Также элементы суперукладки являются одной из основных проблем при предсказании функций белков на основе определения и анализа способов укладки, поскольку белки, обладающие схожими элементами суперукладки, далеко не всегда обладают схожими функциями. Существование таких способов укладки и их широкое распространение среди белковых молекул вынуждает исследователей с осторожностью использовать информацию о связях между известными способами укладки при предсказании функций белков.
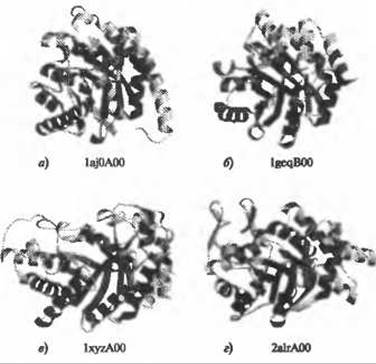
Рис. 6.1. (Цветную версию рисунка см. на вклейке.) Суперучастки укладки, напоминающей структуру бочонка TIM (ß/a)8. Схематичные изображения четырех белков, имеющих укладку бочонка (ß/a)8, представленные в различных надсемействах САТН (и SCOP): а) дегидроптероатсинтаза Е. coli (идентификационный номер (ИН) домена в САТН: 1aj0A00); б) альфа-субъединица триптофан-синтазы Р. furiosus (ИН домена в САТН: IgeqBOO); в) эндо-1,4-бета-ксиланаза Z С. thermocellum (ИН домена в САТН: IxyzAOO); г) альдегидредуктаза Н. sapiens (ИН домена в САТН: 2а1rА00). Структуры совмещались с помощью CORA(Orengo 1999). Молекулы ориентированы одинаково. Общие элементы четырех структур показаны красным цветом. Положение каталитических остатков (согласно Атласу каталитических центров) показано зеленым цветом. Несмотря на значительные структурные различия и отсутствие доказательств гомологии между этими белками, каталитические центры всегда находятся вблизи С-конца центральных ß-тяжей. Изображения пространственных структур были созданы с помощью программы Molscript (Kraulis 1991), рендеринг изображений осуществлялся с помощью программы Raster3D (Merritt and Bacon 1997)