БІОХІМІЯ ДЛЯ ВЧИТЕЛЯ - Ф.Ф. БОЄЧКО - 1985
УТВОРЕННЯ ЖИВИХ СИСТЕМ ТА ЇХ СТРУКТУРНА ОРГАНІЗАЦІЯ
ВІД ПРОСТИХ МОЛЕКУЛ — ДО ЖИВИХ ОРГАНІЗМІВ
Живі системи характеризуються специфічними закономірностями та особливостями структури й функцій, що виникли в процесі тривалого розвитку від простих молекул до високоорганізованих, спеціалізованих і впорядкованих систем.
За визначенням Ф. Енгельса, «життя є спосіб існування білкових тіл, істотним моментом якого є постійний обмін речовин із зовнішньою природою, що їх оточує, причому з припиненням цього обміну речовин припиняється і життя, що приводить до розкладу білка». На основі сучасних наукових досягнень у галузі біологічних наук положення Енгельса про суть життєвих явищ було підтверджено і розвинуто далі.
Радянський учений О. І. Опарін у 1924 р. розробив теорію походження життя на Землі, яка була підтримана вченими різних країн і нині є найбільш повною та науково обгрунтованою. Згідно з цією теорією виникнення життя на Землі є невід’ємною складовою частиною загального еволюційного розвитку матеріального світу. Це тривалий однобічно спрямований процес поступового ускладнення органічних речовин і відповідних цілісних систем, які з них сформувалися, перебуваючи в постійній взаємодії з навколишнім середовищем.
О. І. Опарін докладно розкрив шляхи первинного виникнення органічних речовин з неорганічних молекул і процеси подальшого перетворення їх в білкові тіла, що мають характерні ознаки живого. Весь процес перетворення неживої матерії в живу, за теорією О. І. Опаріна, складається з трьох послідовно спрямованих взаємозумовлених етапів: абіогенний синтез органічних молекул з неорганічних →концентрування органічних речовин в мікроскопічні багатомолекулярні системи (макромолекули)→вдосконалення надмолекулярних білково-ліпоїдно-полінуклеотидних комплексів (коацерватів).
На мал. 1 подано загальну схему основних етапів виникнення життя на Землі та процесів, що їх супроводжували. Як видно зі схеми, основними елементами, сполуки яких стали вихідним матеріалом, необхідним для синтезу органічних сполук, є вуглець, кисень, водень, азот та фосфор. Це так звані біогенні елементи, які становлять основну масу (99 %) органічної речовини живих організмів. З участю цих елементів утворилась велика кількість сполук, з яких в результаті послідовного ускладнення виникли органічні речовини, що дали початок складним надмолекулярним структурам і системам.
Найпростішими представниками органічних сполук е вуглеводні — сполуки вуглецю і водню. За даними сучасної планетної космогонії, вуглеводні досить розповсюджені: їх знайдено на поверхнях планет, у міжпланетній газово-пиловій матерії, у складі комет та метеоритних тіл.
Методом спектрального аналізу найпростіші сполуки вуглецю з воднем (СН2, СН4), а також з іншими елементами виявлено в атмосфері Сонця, температура поверхні якого 5000—7000 °С. Численні факти щодо можливості абіогенного походження органічних речовин дістали також на основі вивчення метеоритних тіл, які потрапляють на Землю. Крім залізо-нікелевих метеоритів, дуже часто зустрічаються кам’янисті метеорити. У їх складі переважають силікати, оксиди різних металів (Mg, AI, Са, Na). Серед кам’янистих метеоритів особливої уваги заслуговують вуглисті хондрити, які містять значну кількість вуглецю (до 5 %) у вигляді графіту та різних органічних сполук — низькомолекулярних вуглеводнів та полімерних сполук аліфатичного ряду, що подібні до сполук, які зустрічаються в сучасних живих організмах. Деякі вчені вважають, що синтез органічних молекул та їх високомолекулярних полімерів відбувався безпосередньо на частинках космічного пилу, покритих льодом і конденсованими газами, під впливом сонячного випромінювання. В результаті цих процесів утворюються хондритні тіла з високим вмістом вуглецю. Оскільки, за найпоширенішою гіпотезою, наша планета утворилася при відносно низьких температурах внаслідок конденсації неоднорідних за складом частинок міжпланетної речовини, серед якої велику кількість становили скупчення хондритних тіл, деякі вчені припускають, що саме ці готові органічні речовини були основою для наступного розвитку життя. Проте готові органічні речовини, що утворились до формування планети, не могли відіграти важливої ролі у виникненні життя, оскільки внаслідок поступового локального підігрівання її за рахунок теплоти, що виділялась під час розпаду радіоактивних елементів, значна частина органічних речовин розпалася б внаслідок піролізу. Тому правильним е твердження, що основна маса органічних речовин утворилася на поверхні Землі з вуглецевих сполук, що виділились у первинну атмосферу при формуванні земної кори — літосфери внаслідок переходу із зовнішніх шарів Землі найбільш легкоплавких сполук.
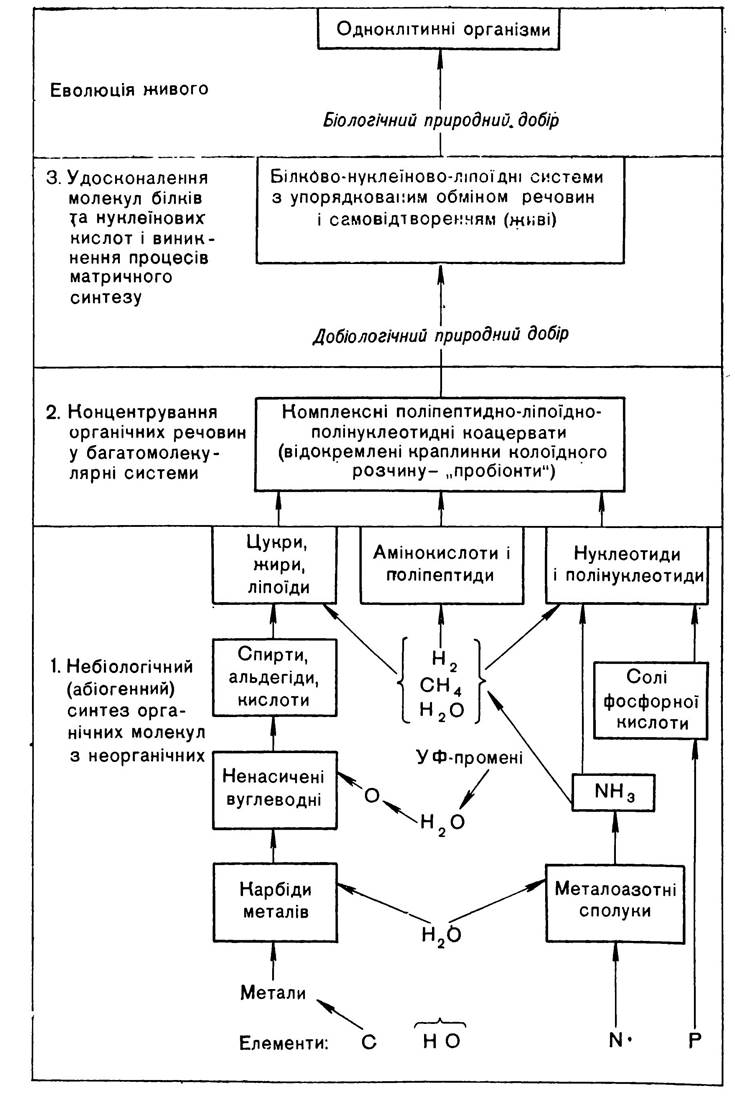
Мал. 1. Ступені і основні процеси виникнення життя на Землі (За П. І. Гупалом).
З процесами, які відбувалися в літосфері, тісно пов’язані процеси утворення гідро- і атмосфери нашої планети. Відомо, що первинна міжпланетна речовина, з якої утворилась наша планета, містила в своєму складі 76 % водню. Частина його витратилась у процесі утворення різноманітних сполук і виділилась при формуванні літосфери, внаслідок чого утворилась земна атмосфера з різко вираженими відновними властивостями.
Під час формування літосфери утворювались також і виділялись інші органогенні елементи. Так, від взаємодії нітридів з гідратною водою земних надр утворювався і виділявся аміак. Сполуки сірки з металами стали джерелом сірководню. Відносно кисню, вважають, що він у вільному стані був присутній у первинній атмосфері Землі лише в мізерних кількостях. Основна маса кисню сучасної атмосфери, як і азоту, виникла значно пізніше біогенним шляхом, в результаті життєдіяльності організмів. Кількість води на поверхні Землі в той час становила 10 % сучасного вмісту її в морях і океанах, решта її утворилась значно пізніше.
Після докладного аналізу складу метеоритів було встановлено, що вуглець в них міститься у вигляді сполук з металами— карбідів та графіту. Природні карбіди (мінерал когеніт)—сполуки вуглецю із залізом, кобальтом, нікелем становлять значну частину матеріалу метеоритних тіл.
Когеніти виявлено також у глибинних породах земної кори. Внаслідок взаємодії цих сполук з водою утворювались різні вуглеводні, насамперед метан, який нагромаджувався в первинній атмосфері Землі.
Вважають, що вуглеводні могли утворюватись і іншими шляхами — прямим відновленням графіту вільним воднем, що виникав внаслідок радіохімічного розпаду води, а також внаслідок піролізу первинних органічних речовин, які входили до складу міжзоряної речовини. Крім вуглеводнів, у первинній атмосфері Землі нагромаджувалась велика кількість аміаку, водяної пари, ціаніду водню (HCN), вуглекислого газу та інших сполук, а також вільних радикалів, які активно вступали у взаємодію одні з одними. В результаті цих реакцій утворювались органічні речовини і насамперед вуглеводні. Ці процеси відбувались протягом перших двох мільярдів років. Підтвердженням абіогенного походження органічних сполук є дані, здобуті в процесі геохімічних і радіоастрономічних досліджень. Було виявлено, що й тепер у глибоких шарах літосфери, де немає ніяких ознак життя, відбуваються процеси абіогенного утворення вуглеводнів. Так було відкрито газові й нафтові родовища в тих шарах земних надр, які не мають безпосереднього зв’язку з осадовими породами. Тому і вважають, що нафта буває як біогенного, так і абіогенного походження. За даними радіоастрономії та планетної космогонії, у міжзоряному просторі та в атмосфері багатьох планет виявлено аміак, водяну пару, метан і навіть ціанід водню та формальдегід, які виникли абіогенним шляхом внаслідок реакцій, що супроводжували формування планет. Усі ці дані є незаперечним підтвердженням того, що на Землі, як і на інших планетах, відбувалось абіогенне утворення вуглеводнів та їх похідних. Слід наголосити, що утворення та дальша хімічна еволюція цих сполук відбувались в первинній атмосфері і на поверхні Землі в умовах, що відрізняються від сучасних.
Дослідник М. Руттен зазначає, що відносно земної поверхні треба чітко розрізняти два періоди — подібний до сучасного — актуалістичний та доактуалістичний, що відрізняється від нього і передував йому. Особливістю доактуалістичного періоду було те, що атмосфера нашої планети мала різко виражений відновлювальний характер. Оскільки не було вільного кисню, не могло відбуватись пряме окислення вуглеводнів та не міг формуватись у верхніх шарах атмосфери озоновий екран — шар газу озону (Оз), який поглинає короткохвильові ультрафіолетові промені, що згубно діють на все живе. Крім того, у цей період над поверхнею Землі гриміли грози, атмосферу пронизували сильні електричні розряди, ішли зливи. Короткохвильове ультрафіолетове та радіоактивне випромінювання, іскрові розряди, висока температура розжареної лави, що виривалась на поверхню Землі через тріщини й розломи, були важливим джерелом енергії для забезпечення перебігу хімічних реакцій і хімічної еволюції утворених сполук. Нині на земній поверхні таких умов немає. Озоновий шар, що міститься на висоті 30 км, надійно захищає нашу планету від згубної сонячної радіації; сучасна атмосфера, поверхні літосфери та гідросфери мають окислювальний характер. Живі організми, які населяють планету, споживають органічні речовини, що перешкоджає їх тривалій еволюції. Тому, за висловом О. І. Опаріна, життя не може виникнути в сучасних умовах на нашій планеті саме тому, що воно вже виникло.
З виникненням життя на земній поверхні відбулась така зміна умов, яка виключає можливість тривалої еволюції органічних речовин таким шляхом, яким вона відбувалась у доактуалістичний період існування нашої планети. Відтворити умови, які були на Землі в період, що передував виникненню життя, можна лише у наукових лабораторіях. Американський біохімік С. Міллер в 1957 р. продемонстрував учасникам міжнародного симпозіуму з питань походження життя утворення амінокислот з метану, водню, аміаку і водяної пари пропусканням електричних розрядів крізь суміш цих газів у спеціально сконструйованому приладі.
У цей самий час подібний синтез здійснили також радянські вчені. Склад суміші в них був дещо інший: замість водню вони брали оксид вуглецю (II).
Згодом у лабораторних умовах, подібних до тих, що були на Землі в перші мільярди років, було здійснено синтез' поліпептидів, полінуклеотидів та інших сполук. З участю поліфосфатів, використовуючи короткохвильове ультрафіолетове випромінювання, здійснено абіогенний синтез АТФ — важливої макроергічної сполуки всіх живих організмів. Таким чином, експериментально було доведено, що в певний період розвитку Землі в гідросфері нагромаджувались неорганічні солі та органічні сполуки різного ступеня складності, що могли вступати в хімічну взаємодію. Підраховано, що за один мільярд років масова частка абіогенно утворених органічних сполук у водах Світового океану могла досягти 1 %. Особливо висока концентрація органічних речовин могла бути в окремих ізольованих водоймах за рахунок випаровування води.
Крім того, постійно відбувались не тільки кількісні, а й якісні зміни, що підпорядковувались в основному загальним фізико-хімічним законам, тобто вирішальним фактором розвитку в той час була фізико-хімічна еволюція різноманітних органічних речовин.
Біологічна еволюція органічних сполук звичайно не була цілеспрямованою, тому на даному етапі в хаосі безконтрольних хімічних реакцій не могли утворюватись речовини з впорядкованою внутрімолекулярною структурою і певними специфічними властивостями та функціями. Утворення їх стало можливим лише тоді, коли внаслідок тривалого розвитку сформувались певні ізольовані вихідні системи, в яких виник певний порядок взаємодії із зовнішнім середовищем. Саме в процесі взаємодії та пов’язаного з нею відбору індивідуальних вихідних систем внутрішня організація цих систем пристосовувалась до зовнішніх умов, відбувалась більш досконала взаємодія речовин і процесів, що привело до закономірного організованого обміну речовин. Виникнення високомолекулярних комплексних систем становить другий, вищий ступінь еволюції органічних речовин.
У 60-х роках XX ст. американський дослідник С. Фокс експериментально довів можливість синтезу високомолекулярних систем із застосуванням термічного впливу на суміш амінокислот. Утворені частинки осаджувались з 1-процент- ного водного розчину хлориду натрію у вигляді мікроскопічних кульок, внаслідок чого вони дістали назву мікро- сфер.
Щодо способів утворення високомолекулярних комплексних систем (коацерватів) та виділення їх з однорідного первинного розчину органічних речовин є багато теорій. Найбільш поширена коацерватна теорія утворення багато- молекулярних систем, яку сформулював О. І. Опарін. Коацервати (від латинського coacervo — нагромаджувати, збирати до купи) можна легко добути в лабораторних умовах внаслідок змішування розчинів як природних (білків, нуклеїнових кислот), так і штучних полімерів. На думку вчених, такі коацерватні частинки у природних умовах доактуалістичного періоду утворювались поступово. На першому його етапі молекули різних речовин концентрувались і зближувались у певних місцях «первинного бульйону». Зближені молекули об’єднувались, утворюючи значні скупчення їх. Після досягнення певних розмірів водні оболонки молекул змінювалися, утворюючи мікроскопічні краплини, оточені спільною оболонкою,— коацервати (мал. 2).
Коацерватні краплі можуть переміщуватися в рідині, вибірково адсорбувати невеликі молекули з навколишнього середовища. При дослідженні їх властивостей було виявлено, що в коацерватних краплях можуть відбуватись впорядковані хімічні процеси, особливо коли до їх складу добавити активні біокаталізатори. О. І. Опарін в коацерватні краплі, які складалися з поліглікозиду і гістону, добавляв відповідні ферменти, з участю яких при наявності в навколишньому середовищі глюкозо-1-фосфату відбувався синтез крохмалю. Об’єм краалі при цьому значно збільшувався. За ЗО хв вона зростала в 1,5 раза. При добавлянні краплі ферменту, який сприяє гідролітичному розщепленню полісахаридів, утворювалась мальтоза, і розміри краплі значно зменшувались. Підбираючи відповідні ферменти, учений проводив синтез або розкладав полінуклеотиди, що входили до складу коацерватних крапель. Ці дослідження свідчать про те, що утворення коацерватів у водоймах в доактуалістичний період було новим, вищим етапом еволюції абіогенно утворених органічних сполук. Якщо до того часу органічні речовини вільно «плавали» в навколишньому середовищі, з яким вони становили єдине ціле, то після утворення коацерватів молекули органічних речовин відокремились від нього і створили певні цілісні індивідуальні високомолекулярні системи, що могли вступати в певні примітивні обмінні реакції. Маючи здатність до вибіркової адсорбції речовин з навколишнього середовища, різні коацерватні краплі значно відрізнялися між собою іонним складом, якістю та кількістю полімерних сполук, внутрішньою структурою та мали інші індивідуальні особливості. Тому дальший розвиток таких систем залежав уже не тільки від умов навколишнього середовища, а й від їх внутрішньої специфічної організації. Створилась певна залежність між індивідуальною організацією системи і характером тих перетворень, які в ній відбувались. Тому в цих умовах тривалий час могли існувати лише ті системи, індивідуальна організація яких забезпечувала сприятливий для їх існування перебіг хімічних реакцій і процесів.
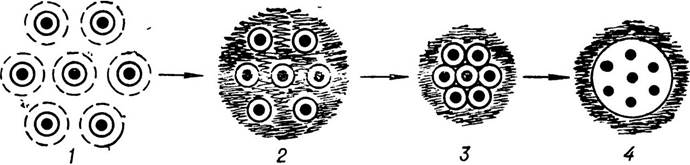
Мал. 2. Схема утворення краплин коацервату:
1 — молекули білка, оточені шарами молекул води; 2, 3 — зменшення водних оболонок у молекул білка і їх зближення;
4 — утворення коацервату.
Отже, на даному етапі розвитку організованих багатомолекулярних систем виник свого роду добіологічний добір, під контролем якого відбувалась їх дальша еволюція, що набула певної спрямованості. Внаслідок такої спрямованої еволюції індивідуальні коацерватні краплі набували характеру динамічних відкритих систем, що могли самозберігатись і тривалий час існувати завдяки постійній взаємодії із зовнішнім середовищем, здатності до росту і збільшення маси за рахунок навколишнього середовища та здатності до розмноження, поділу не лише під впливом зовнішніх факторів, й за рахунок внутрішніх процесів. Таким чином, внаслідок взаємодії вихідних систем з навколишнім середовищем постійно збільшувалась кількість цих систем. Системи ставали не тільки динамічно стійкими, а й більш швидкими. Більшу можливість зберігатись і розвиватись мали ті системи, які набули здатності швидко перетворювати речовини, тобто були більш динамічними, і водночас могли здійснювати реакції, які б не порушували динамічної рівноваги системи. Такі системи О. І. Опарін називає протобіонтами, попередниками живого, наголошуючи цим, що, незважаючи на складнішу і досконалішу організацію їх порівняно із статичними коацерватами, ці системи ніяк не можна віднести до живих істот (еубіонтів). Тільки в результаті наступної тривалої еволюції та добіологічного природного добору протобіонти могли перетворитися в первинні живі організми, які дали початок життю на Землі.
Вдосконалення комплексних динамічних систем (протобіонтів), яке відбувалось протягом сотень мільйонів років під впливом добіологічного природного добору, за теорією О. І. Опаріна, є останнім ступенем виникнення життя на Землі. Деякі вчені заперечували можливість природного добору в комплексних багатомолекулярних системах та протобіонтах, вважаючи, що він властивий лише живим організмам. На думку О. І. Опаріна, виникнення життя не можна звести лише до дії законів неорганічної природи, тому становлення природного добору передує виникненню життя. Такий добіологічний природний добір сприяв тому, що будь- яке порушення узгодженості процесів всередині системи призводило до загибелі, зникнення її. І навпаки, підвищення узгодженості та ефективності процесів зумовлювало збереження і розмноження досконаліших систем.
Отже, на основі добіологічного природного добору постійно вдосконалювались організації значної кількості про- тобіонтів. На початкових етапах еволюції протобіонтів коацерватні краплі збільшувалися завдяки процесам полімеризації складових компонентів при наявності в навколишньому середовищі багатих на енергію фосфатів, що утворилися абіогенно в первинному середовищі за рахунок енергії короткохвильового випромінювання. Проте з часом внаслідок збільшення кількості коацерватів та інтенсивного використання макроергічних сполук в більш сприятливих умовах виявилися ті коацерватні краплі, в яких у процесі еволюції виробилась здатність самостійно забезпечувати себе енертією за рахунок певних внутрішніх процесів, насамперед окислювально-відновних реакцій.
Але забезпечити такі складні процеси прості каталізатори, що перебували в навколишньому середовищі,— іони металів Zn, Cu, Fe були неспроможними. Тому для дальшого розвитку протобіонтів важливе значення мало вдосконалення процесів каталізу — основного фактора організації обміну речовин. Таке вдосконалення відбувалось у процесі еволюційного розвитку протобіонтів на основі природного добору, внаслідок включення каталізаторів до надмолекулярної системи протобіонтів і закріплення в ній. Вибірково вбираючись з навколишнього середовища, окремі складові частини каталітично активної молекули створювали всередині протобіонту певний каталітично активний комплекс. Якщо цей комплекс позитивно впливав на певні реакції полімеризації чи інші процеси, які зумовлювали динамічну стійкість системи та здатність її до росту, кількість таких систем значно збільшувалась і займала чільне місце в еволюції протобіонтів. Системи, які мали менш досконалі механізми каталізу, поступово зникали. Таким чином, з численних комбінацій здатних до каталізу молекулярних угруповань відбирались лише найефективніші, які згодом повинні були стати універсальними для живих істот. Такою універсальною сполукою, що забезпечує процеси оксидоредукції в усіх без винятку живих організмах, є складні ферменти, у яких нікотинамідаденіндинуклеотид (НАД) — активна група. Ці ферменти забезпечують складний процес перенесення водню по системі дихального ланцюга, окислення органічних сполук та синтез макроергів — багатих на енергію нуклеотидів (АТФ). Універсальність цих ферментів для всіх живих систем свідчить, що саме НАД був відібраний живою природою з усієї кількості аналогічних сполук ще на ранніх етапах розвитку в процесі вдосконалення протобіонтів. Отже, в ході добіологічного добору поступово відбувалось формування складної системи ферментативних процесів, в яких основну роль відігравали коферменти, що були більш досконалими порівняно з неорганічними каталізаторами.
Здійснення синтезу коферментів було важливим кроком вперед на шляху еволюції протобіонтів. Таким чином створились умови для існування протобіонтів у середовищі певного складу. Усе це потребувало узгодженого перебігу великої кількості реакцій, необхідних для синтезу. Тому одночасно з удосконаленням процесів каталізу в результаті добіологічного прйродноґо Добору протобіонтів вдосконалю вались та ускладнювались обмінні реакції: збільшувалась їх кількість, подовжувались утворені ними ланцюги, розгалужувались і замикались цикли.
Якщо вихідні форми протобіонтів були повністю гетеротрофні і залежали від складу навколишнього середовища, то внаслідок наступної еволюції утворились системи із значною кількістю комплексних багатоланцюгових процесів, які зменшували залежність протобіонтів від зовнішнього середовища. З ускладненням системи обмінних реакцій виникла потреба в нових, більш досконалих порівняно з коферментами, високоспецифічних каталізаторах, внутрімолекулярна структура яких була б добре пристосована до виконуваних ними функцій. Оскільки всі ферменти мають білкову природу, тобто є поліпептидами з певною, характерною для кожного з них первинною структурою, синтез їх міг здійснюватись лише при наявності досить складних механізмів, які б забезпечували строго закономірну побудову всієї молекули. Причому ця закономірність стосується не тільки точної послідовності амінокислотних залишків у по- ліпептидному ланцюгу, а й просторового розміщення, упаковки всієї молекули в глобулі, бо це є важливою умовою забезпечення їх каталітичної активності. Іншими словами, для наступної прогресивної еволюції протобіонтів важливе значення мало виникнення білоксинтезуючого апарату, здатного до синтезу білків із строго специфічною первинною структурою. Немає сумніву в тому, що складний і досконалий рибосомально-матричний, білоксинтезуючий апарат сучасних організмів був сформований в результаті довготривалого еволюційного розвитку протобіонтів і є його вищим етапом. Шляхи і напрями добіологічної еволюції цьогб( апарату поки що досить гіпотетичні. Вважають, що внаслідок еволюції та добіологічного добору в протобіонтах значно посилилась полімеризація пептидів і нуклеотидів. Цьому особливо сприяли виникнення спряжених енергетичних реакцій і наявність неорганічних каталізаторів та коферментів. Поступово почала виявлятися певна взаємодія між молекулами різних полімерів, створилася система, в якій полімеризація молекул нуклеотидів була пов’язана з формуванням певної послідовності амінокислот — первинної структури поліпептидів. Зрозуміти це можна на прикладі деяких сучасних мікроорганізмів, в яких синтез поліпептидів відбувається поза рибосомально-матричним апаратом, з участю нуклеозидфосфатів. Схема цього процесу така: нуклеозидтрифосфат + амінокислота → нуклозиддифосфат+пептид+ортофосфат.
При цьому для кожного з чотирьох нуклеозидфосфатів (АТФ, ГТФ, ЦТФ, УТФ) існує певна група амінокислот, яку вони можуть вибірково приєднувати до другого вуглецевого атома рибози.
Якщо комбінація амінокислотних залишків, що виникла під впливом полінуклеотиду, сприяла перебігу в протобіон- ті реакцій, необхідних для його існування, то протобіонт діставав перевагу над іншими аналогічними системами в швидкості свого росту і розвитку. У протилежному випадку дана комбінація амінокислот разом з протобіонтом, в якому вона виникла, зникала під впливом природного добору. Так відбувався природний добір не окремих полінуклеоти- дів чи поліпептидів, а цілісних систем — протобіонтів, які відповідали даним умовам. Полінуклеотиди при цьому відігравали важливу роль стабілізуючих факторів еволюції. У них відбувалось просторове закріплення певного порядку чергування мононуклеотидних ланок, з участю яких відбувався синтез каталітично вигідних амінокислотних комбінацій.
На наступному етапі вдосконалення білоксинтезуючого апарату функції полінуклеотидів поступово ускладнювалися і видозмінювалися, внаслідок чого виникли різні види полінуклеотидів — ДНК і РНК. Вважають, що спочатку здатність до збереження і передачі інформації при синтезі поліпептидів була властива РНК-подібним полінуклеоти- дам. Наступний поділ функцій відбувся між двома видами нуклеїнових кислот — дволанцюговою ДНК і одноланцюговою РНК, з яких перша менш метаболічно активна, вона «спеціалізувалась» на самовідтворенні і набула здатності до реплікації, а друга — на безпосередній участі в синтезі білка. Це було прогресивним кроком у процесі розвитку. З виникненням здатних на самовідтворення складних надмолекулярних відкритих систем відбувається перехід від неживої матерії до якісно нової форми її існування, яку ми називаємо життям.
На даному етапі внаслідок хімічної еволюції з неживих молекул утворились найпростіші примітивні живі організми, що мали всі характерні особливості живих істот, якими вони відрізняються від неживої природи. Це здатність до вибіркового вбирання речовин з навколишнього середовища та екскреції продуктів обміну, здатність до росту, розвитку, розмноження і самовідтворення, руху та подразливості. Внаслідок наступної біологічної еволюції виникла вся різноманітність живих організмів, які населяють нашу планету.
Сучасна наука дедалі глибше проникає в таємниці життя, пізнає шляхи його виникнення та розвитку від неорганічних молекул до живих систем. Значний вклад у розв’язання цієї проблеми внесли радянські вчені. У нашій країні широко проводяться експериментальні дослідження, на основі яких дедалі чіткіше вимальовуються реальні шляхи виникнення та розвитку життя на нашій планеті. Розроблена О. І. Опаріним теорія перетворення неживої матерії в процесі складної хімічної і біологічної еволюції в якісно нову, вищу форму існування матерії, має не тільки світоглядне значення, а й свідчить про перемогу діалектико-матеріалістичного світорозуміння над релігійним та ідеалістичним. Важливою є ця теорія для розвитку біологічної науки і природознавства в цілому, оскільки з’ясування таємниці виникнення та розвитку життя — необхідна умова пізнання його суті. А це в свою чергу, відкриває широкі можливості для використання досягнень сучасної науки в керуванні процесами життєдіяльності в інтересах людства. Питання про походження життя тісно пов’язане з питанням про його суть, що є однією з найважливіших проблем біологічної науки. Розв’язання цієї проблеми забезпечить небачений прогрес у розвитку людства, дасть людині значну владу над природою.