БІОХІМІЯ ДЛЯ ВЧИТЕЛЯ - Ф.Ф. БОЄЧКО - 1985
ЕНЕРГЕТИКА БІОЛОГІЧНИХ ПРОЦЕСІВ
ФОТОСИНТЕЗ — ОСНОВНЕ ДЖЕРЕЛО СИНТЕЗУ ОРГАНІЧНИХ РЕЧОВИН
Фотосинтез — надзвичайно важливий і специфічний процес, що відбувається в біосфері. Він виник на певному етапі розвитку живої матерії внаслідок вдосконалення примітивних форм фіксації оксиду вуглецю (IV). Цей унікальний процес становить одну з основних ланок біологічного кругообігу речовин і є основою життя на нашій планеті.
Під впливом енергії сонячного випромінювання в зелених рослинах здійснюється синтез органічних речовин з неорганічних і відбувається збагачення атмосфери киснем. Потенціальна енергія готових органічних речовин — продуктів фотосинтезу, використовується тваринними організмами для забезпечення численних метаболічних процесів. Таким чином, рослини є ніби посередниками між Сонцем ї всім живим на Землі. У цьому полягає важлива космічна роль зелених рослин. Жодний процес, який відбувається на нашій планеті, не заслуговує на таку увагу, як процес, що відбувається в зелених листках, коли на них падає сонячний промінь.
Загальна схема фотосинтезу має такий вигляд:
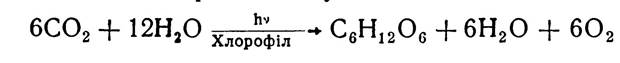
Цей процес забезпечує багатоступеневе перетворення світлового електромагнітного випромінювання певної довжини хвилі в хімічну енергію органічних сполук і здійснюється з участю складних біологічних систем і структур рослинної клітини. Відбувається фотосинтез в специфічних органоїдах рослинних клітин — хлоропластах. Як зазначалося, хлоропласти мають гетерогенну гранулярно-ламелярну структуру і перебувають у постійному взаємозв’язку з цитоплазмою. Перші уявлення про структуру хлоропластів і функції хлорофілу були сформульовані ще в 1903 р. російським фізіологом К. А. Тімірязєвим. Пізніше з’ясували окремі ланки цього складного процесу. Проте й зараз механізм фотосинтезу до кінця не розгадано.
Останнім часом завдяки електронномікроскопічним дослідженням встановлено особливості фотосинтетичних одиниць (квантосом), розміщених в окремих ділянках хлоропластів, що утворені плоскими мішкоподібними структурами — тилакоїдами. Кожний тилакоїд має форму диска, в якому щільно упаковані фотосинтетичні одиниці, що містять 200—250 молекул хлорофілу та інших пігментів, зв’язаних з білково-ліпідними асоціаціями.
Фотосинтетична активність властива не всім молекулам пігментів, які входять до складу квантосом. Поглинання кванту світла відбувається послідовно кожною з цих молекул, у середньому один раз за 0,1 с. Енергія всіх квантів концентрується в фотохімічно активному ферментативному центрі, де з участю складних біохімічних процесів відбувається перетворення її в хімічну енергію органічних сполук.
У дослідження структури і властивостей хлорофілу значний внесок зробили російські та зарубіжні вчені М. С. Цвєт, К. А. Тімірязєв, М. В. Ненцький, Р. Вудворд, Г. Фішер та ін.
У вищих рослин та фотосинтезуючих водоростей існує два види хлорофілу — хлорофіл a (C55H72О6N4Mg) і хлорофіл в (C55H70О6N4Mg). Структурна формула хлорофілу а має такий вигляд:
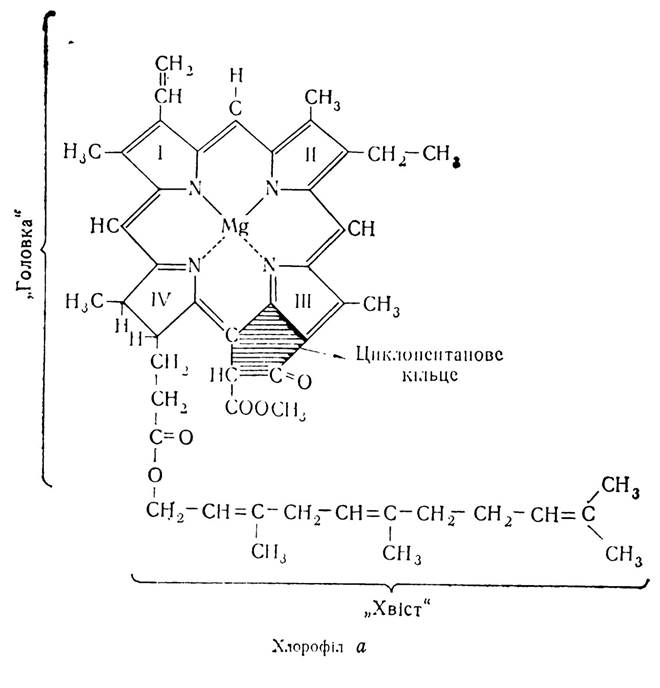
Як видно з формули, до складу хлорофілу входить порфіринове ядро, що складається з чотирьох пірольних залишків, зв’язаних за допомогою двох основних і двох додаткових валентних зв’язків з атомом магнію. Ця частина молекули утворює гідрофільну головку. Друга частина молекули складається із залишку високомолекулярного ненасиченого спирту фітолу і утворює гідрофобний хвіст. Така будова хлорофілу відіграє важливу роль в фіксації його молекул у ламелярній структурі хлоропластів. Порфіринове ядро молекули розміщується на поверхні мембран під кутом 35—40° до площі шару ламел, а залишки спирту фітолу занурені в ліпідний шар ламелярних мембран.
На відміну від інших порфіринів, хлорофіл а має вищий вміст водню і, крім чотирьох пірольних, має ще й циклопентанове кільце, розміщене біля третього пірольного залишку. Хлорофіл в відрізняється від хлорофілу а деякими деталями в будові, а саме: біля другого пірольного залишку замість метильної групи міститься формільна 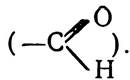
У рослинах хлорофіл в може утворюватись з хлорофілу а. Бактеріохлорофіл відрізняється від хлорофілу а тим, що вінільна група (—СН=СН2) біля першого пірольного кільця замінена ацетильною групою 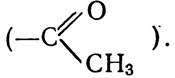
Правильність структури хлорофілу була доведена внаслідок хімічного синтезу його, здійсненого в 1960 р. американським хіміком Р. Вудвордом.
До групи хлорофілів належить також хлорофіл с, який міститься в клітинах бурих водоростей і відрізняється від хлорофілу а і в будовою бокових радикалів біля II і IV пірольних кілець.
Крім хлорофілу, у хлоропластах міститься ще ряд речовин, які беруть безпосередню участь у певних реакціях фотосинтетичного процесу: каротиноїди, пластохінони, пластоціаніни, фередоксин, флавопротеїди та іони металів.
До каротиноїдів належить велика група жовто-оранжевих пігментів, серед яких найважливішими є каротин і ксантофіл. Ці пігменти забезпечують забарвлення плодів, овочів, жовтка яєць, пір’я птахів та ін.
Каротиноїди відомі давно як пігменти, що супроводжують хлорофіл, однак, їх функції в процесі фотосинтезу стали відомі лише в останні десятиріччя. В результаті досліджень встановлено, що не всі пігменти рослин відіграють у процесі фотосинтезу однакову роль. Основним пігментом фотосинтезу вищих рослин є хлорофіл а, що міститься в усіх здатних до фотосинтезу клітинах. Хлорофіл в і каротиноїди є додатковими пігментами, які разом з основним у структурі хлоропластів створюють комплексні фотосинтетичні системи, які забезпечують інтенсивне використання спектра сонячного випромінювання. Кожний з цих пігментів має здатність поглинати світлові хвилі певної довжини оранжево-червоних і синьо-фіолетових частин спектра. Так, хлорофіл а поглинає світлові хвилі довжиною 680—700 нм, а хлорофіл в — 650 нм. Крім того, виявлено, що хлорофіл а присутній у клітині у вигляді кількох форм, які мають різні спектри поглинання і різні фотохімічні функції.
Спільне функціонування ряду пігментів з різними спектрами поглинання забезпечує засвоєння ними досить широкого спектра електромагнітного випромінювання. Завдяки адсорбційним властивостям всі пігменти мають здатність поглинати світлову енергію, та лише незначна частина їх використовується для перетворення цієї енергії в хімічну. Цей процес забезпечується лише спеціалізованими фотохімічно активними пігментами, а інші пігменти виконують роль акумуляторів квантів світла, тобто є додатковими, допоміжними. Встановлено, що допоміжні пігменти можуть передавати до 80—100 % ввібраної енергії на основні пігменти, внаслідок чого ефективність фотосинтезу значно підвищується. Це явище називається ефектом Емерсона. Воно було виявлено внаслідок численних спостережень, які свідчили, що ефективність фотосинтезу значно збільшується внаслідок освітлення рослин променями монохроматичного світла з різною довжиною хвиль. Так було встановлено, що інтенсивність фотосинтезу, який спостерігається при довжині хвилі 700 нм, значно посилюється при додатковому освітленні світлом довжиною хвилі 650 нм. Цей ефект посилення пояснюється існуванням у рослинних клітинах двох фотосинтетичних пігментних систем (І і II).
У першій системі основним пігментом з фотосинтетичною активністю (реактивним центром) є хлорофіл а700 (або Р700). Крім того, до складу цієї системи входить ще ряд допоміжних пігментів: а600, а670, а685, а695 хлорофіл в і каротиноїди, цитохром f. У середньому на 450 молекул пігментів припадає 1 молекула хлорофілу а700.
Друга фотосинтезуюча система у вигляді реактивного центру містить хлорофіл а690 а також ряд допоміжних пігментів, які концентрують і транспортують світлову енергію — хлорофіл а678, хлорофіл в, фікобіліни, каротиноїди.
Механізм фотосинтезу
За сучасними уявленнями, фотосинтез — це складна система окислювально-відновних реакцій та фотохімічних процесів, які забезпечують засвоєння і перетворення енергії сонячного випромінювання в енергію хімічних зв’язків АТФ та утворення відновника з високим потенціалом (НАД·Н+Н+) в реакціях фосфорилювання. Складається цей процес з трьох послідовних етапів, які включають світлові і темнові фази. Тривалість перших становить 1·10-5с, а других — 1 · 10-3с. На першому і другому етапах (світлова фаза) з участю хлорофілу відбуваються поглинання світлової енергії та фотохімічний розклад (фотоліз) води з виділенням молекулярного кисню. Одночасно з цим здійснюються окислювально-відновні реакції, в яких крім хлорофілу, беруть участь оксидоредуктази, що переносять електрони від молекул води на НАДФ+, внаслідок чого відбувається вивільнення енергії і акумулювання її в макроергічних зв’язках АТФ. На третій (темновій) фазі використовуються відновна форма (НАД·Н + Н+) і АТФ для відновлення вуглекислого газу до вуглеводів.
Процес фотосинтезу відбувається за основними правилами фотохімії: лише поглинені промені можуть зумовлювати хімічний ефект, пропорційний кількості сприйнятої енергії. Тому перед виконанням певної роботи світлова енергія повинна бути поглинута, тобто фотохімічний ефект можуть виявляти лише кванти світла, які поглинаються пігментами.
Кожна молекула речовини при фотохімічних реакціях поглинає стільки світла, щоб забезпечити запас енергії, який викликає в молекулі відповідні зміни. Таким чином, кожен поглинений квант світла зумовлює елементарний фотоакт, відповідні зміни в молекулі пігменту. Тому при поглинанні кванту світла молекула пігменту переходить у високоенергетичний, збуджений стан і набуває здатності віддавати і приєднувати електрони. Цей процес зумовлений тим, що в молекулі хлорофілу, як і в інших органічних сполуках з подвійними зв’язками, я-електрони при поглинанні світла можуть переходити на вищий енергетичний рівень — кванти світла вибивають ці електрони з молекули хлорофілу, внаслідок чого він окислюється і перетворюється в позитивно заряджений іон, а електрони переносяться на відповідний акцептор:
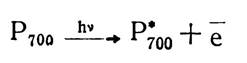
Окислений хлорофіл має здатність приєднувати електрони від донорів, які мають позитивний окислювально-відновний потенціал, і знову набуває здатності поглинати кванти світла. Хлорофіл у цьому процесі виступає як електронна помпа, що перекачує електрони до вищого енергетичного рівня.
У фотосистемі І відбувається перенесення електронів від Е10 (+0,4 В) на Е10(-0,4 В), а в фотосистемі II від Е10(+0,8 В) до Е10 (-0,15 В). У процесі перенесення електронів частина енергії використовується на синтез АТФ з АДФ і неорганічного фосфату. Цей процес називається фотосинтетичним фосфорилюванням. Від окислювального фосфорилювання, яке відбувається в мітохондріях, він відрізняється тим, що відбувається в анаеробних умовах, і при збагаченні середовища киснем зменшується. Крім того, при окислювальному фосфорилюванні в мітохондріях електрони відновлюють кисень, а при фотосинтетичному — інші сполуки. Фотосинтез деякою мірою є процесом, зворотним до біологічного окислення в мітохондріях (тканинного дихання), що видно з наведеної схеми:

При дослідженні фотосинтетичного фосфорилювання було встановлено, що зумовлене енергією світла перенесення електронів може бути циклічним і нециклічним. При циклічному фотосинтетичному фосфорилюванні електрон, вибитий з молекули хлорофілу, витративши додаткову енергію, добуту від кванту світла, повертається до тієї самої молекули хлорофілу, тобто відновлення молекули хлорофілу здійснюється за рахунок повернення його власних електронів. Це так званий круговий потік електронів, що відбувається в електронно-транспортних системах тилакоїдних мембран з участю цитохрому в6 і f. Електрони перекачуються «вгору» з участю квантів світла, а при «спуску» їх, за рахунок енергії активованого світлом хлорофілу, відбувається синтез однієї молекули АТФ. Процес циклічного фосфорилювання проходить на різних ділянках тилакоїдів у гранах та між ними.
При нециклічному фосфорилюванні електрон, вибитий з молекули хлорофілу, використовується на відновлення НАДФ+ до НАДФ·Н + Н+. Відновлення хлорофілу здійснюється за рахунок електронів гідроксид- іонів води, які вивільнюються при її фотоокисленні. При фотоокисленні води (фотолізі) молекули її розпадаються з утворенням протонів і молекулярного кисню:
![]()
Таким чином, при нециклічному фотопереносі (фосфорилюванні) утворюються два продукти — АТФ і НАДФ·Н+Н+. Вивільнення протонів внаслідок фотолізу води врівноважується використанням їх при утворенні відновленої форми нікотинамідних ферментів. Вважають, що акт фосфорилювання в процесі фотосинтезу відбувається на ділянці між донором електрона (Н2О) і хлорофілом, що збуджений світлом. Кінцевим акцептором водню, утвореного в процесі фотолізу води, е оксид вуглецю (IV).
Нижче наведено схему циклічного і нециклічного фотосинтетичного фосфорилювання (за Арноном):
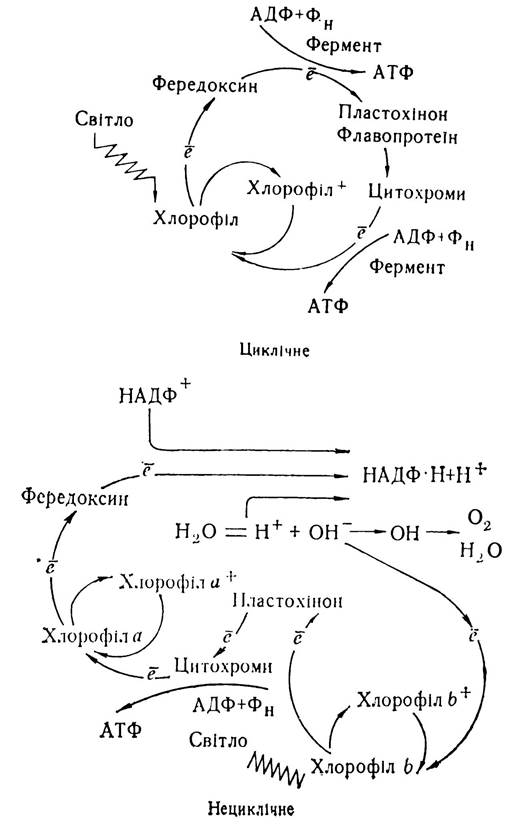
Як видно із схеми, при обох способах фотосинтетичного фосфорилювання електрон від збудженої молекули хлорофілу (х) переноситься на фередоксин, який відновлюється. При нециклічному фосфорилюванні електрон від фередоксину через проміжні темнові реакції переходить разом з іонами водню на молекулу НАДФ+, яка відновлюється до НАДФ·Н+Н+.
Таким чином, у циклі світлових реакцій фотосинтезу відбувається індуковане світлом перенесення електронів, що пов’язане з процесами фосфорилювання і утворенням АТФ та відновленої форми НАДФ. В одному циклі світлової фази утворюється три молекули АТФ і дві молекули НАДФ·Н+Н+, які використовуються в темновій фазі фотосинтезу для відновлення СО2.
За хемосинтетичною теорією Мітчелла, синтез АТФ у світловій фазі фотосинтезу, як і при окислювальному фосфорилюванні в мітохондріях, відбувається з участю протонного насоса мембран тилакоїдів. Пластохінон, фередоксин і НАДФ+ переносять не лише електрони, а й протони, які вивільнюються при фотоокисленні води. Окислювально-відновні системи в мембранах тилакоїдів розміщені, очевидно, так, що протони використовуються на зовнішній мембрані тилакоїдів, а вивільнюються всередині цих структур. Завдяки цьому створюється своєрідний протонний насос, який приводиться в дію електронами. Градієнт концентрації протонів активує мембранну АТФ-азу, внаслідок чого відбувається синтез АТФ.
Утворена АТФ використовується для енергозабезпечення наступної, темнової фази фотосинтезу, а також для ряду інших метаболічних процесів, що здійснюються в клітині — синтезу білків, нуклеїнових кислот, пігментів. Крім того, енергія АТФ використовується для забезпечення процесів руху, транспорту різних речовин, підтримання іонного балансу.
Наступні темнові реакції фотосинтезу можуть відбуватись лише після завершення світлової фази, внаслідок якої утворюються макроергічні сполуки, енергія яких використовується для відновлення оксиду вуглецю (IV) до вуглеводів. Хлорофіл і світло для темнової фази не потрібні.
Темнова фаза фотосинтезу відбувається в стромі хлоропластів у вигляді послідовних перетворень, що ведуть до утворення кінцевих продуктів фотосинтезу та відтворення початкового акцептора оксиду вуглецю (IV) і каталізується ферментами пластид. Цей процес має циклічний характер і, на честь вченого, який займався його докладним вивченням, називається циклом Кальвіна.
Темнові реакції фотосинтезу починаються з участю АТФ і НАДФ·Н+Н+. На відновлення однієї молекули СО2 до вуглеводу використовується три молекули АТФ і дві молекули НАДФ·Н+Н+.
Як свідчать дослідження Кальвіна, первинним акцептором оксиду вуглецю (IV) є енольна форма рибулозо-1,5-дифосфату, яка утворюється з рибулозо-5-монофосфату з участю АТФ. Внаслідок карбоксилювання рибулозо-1,5-дифосфату та наступного гідролізу проміжної сполуки утворюються дві молекули фосфогліцеринової кислоти, яка з участю АТФ фосфорилюється з утворенням 1,3-дифосфогліцеринової кислоти. Реакцію приєднання СО2 до рибулозо-1,5-дифосфату каталізує фермент а-рибулозо-1,5-дифос-фаткарбоксилаза. Потім 1,3-дифосфогліцеринова кислота відновлюється воднем НАДФН+Н+ з участю АТФ та ферменту фосфогліцеринальдегіддегідрогенази до гліцерин-альдегід-3-монофосфату, частина якого ізомеризується в діоксіацетон-3-монофосфат.
На наступному етапі темнових реакцій фотосинтезу дві утворені фосфотріози перетворюються в результаті зворотних реакцій гліколізу в гексозу та відбувається регенерація первинного акцептора оксиду вуглецю (IV) — рибулозо-5-монофосфату, за допомогою реакцій пентозофосфатного циклу. Спочатку відбувається конденсація гліцериналь-дегід-3-монофосфату та діоксіацетон-3-монофосфату з участю ферменту альдолази з утворенням молекули гексози — фруктозо-1,6-дифосфату:
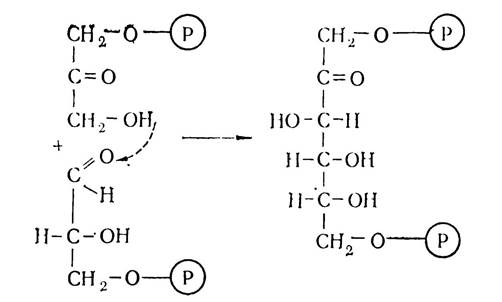
Утворений фруктозо-1,6-дифосфат використовується для синтезу моно- і полісахаридів (глюкози, сахарози, крохмалю), а також дефосфорилюється з утворенням фруктозо-6-монофосфату.
Фруктозо-6-монофосфат може також включатись в пентозофосфатний цикл для регенерації первинного акцептора оксиду вуглецю (IV)—рибулозо-1,5-дифосфату. Здійснюється це двома способами — ізомеризацією фруктозо-6-мо- нофосфату в глюкозо-6-монофосфат, який є початковим субстратом пентозофосфатного циклу, або конденсацією з гліцеринальдегід-3-монофосфатом з утворенням проміжного продукту пентозофосфатного циклу — еритрозо-4-монофосфату. Цей фосфорильований чотиривуглецевий моносахирид може потім конденсуватись з діоксіацетон-3-моно-фосфатом, внаслідок чого утворюється семивуглецева сполука— гептулозо-1,7-дифосфат. Останній втрачає один залишок фосфорної кислоти і перетворюється в гептулозо-7-фосфат, який конденсується з гліцеринальдегід-3-монофос- фатом. При цьому утворюються рибозо-5-монофосфат і ксилулозо-5-монофосфат, які потім ізомеризуються в рибу- лозо-5-монофосфат. Крім того, частина еритрозо-4-фосфату може за допомогою складних перетворень утворювати ряд амінокислот і азотистих основ. Для утворення амінокислот метіоніну, цистину може використовуватись також частина фосфогліцеринової кислоти, яка утворюється на початку циклу Кальвіна. Таким чином, цей цикл є важливим джерелом утворення не лише вуглеводів, а й інших органічних сполук, що використовуються рослинним організмом в найрізноманітніших метаболічних реакціях. Сумарне рівняння реакцій циклу Кальвіна має такий вигляд:
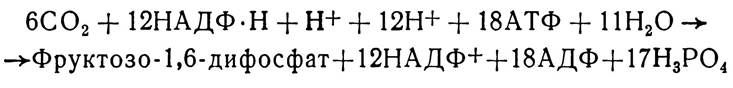
Отже, для синтезу однієї молекули гексози використовується шість молекул СО2. Кожна шоста молекула фосфо-гліцеринового альдегіду виходить з циклу і використовується для синтезу фруктозо-1,6-дифосфату, з якого потім утворюються моно-, ди- і полісахариди. Інші молекули фосфогліцеринового альдегіду з участю АТФ і відповідних ферментів здійснюють регенерацію субстрату СО2 — рибулозо-1,5-дифосфату. Ферменти циклу Кальвіна містяться в стромі, а а-рибулозо-1,5-дифосфаткарбоксилаза — в зовнішній мембрані тилакоїдів.
Крім циклу Кальвіна, існують і інші шляхи засвоєння вуглецю в процесі фотосинтезу. Було встановлено, що акцептором вуглецю може бути не лише рибулозо-1,5-дифосфат, а й тривуглецева сполука — фосфоенолпіровиноградна кислота. При карбоксилюванні останньої з участю ферменту фосфоенолпіруваткарбоксилази утворюється чотири- вуглецева органічна кислота — щавлевооцтова (оксало-ацетат). Остання з участю НАДФ-Н+Н+ відновлюється до малату (яблучної кислоти) або перетворюється в фосфо- гліцеринову кислоту, а потім глюкозу. З малату окислювальним декарбоксилюванням утворюються СО2 і НАДФН+Н+, які можуть використовуватись у циклі Кальвіна. Утворений в цьому процесі піруват фосфорилюється з участью АТФ, внаслідок чого утворюється первинний акцептор СО2 — фосфоенолпіруват.
Рослини, у яких першим продуктом конденсації є фосфогліцеринова кислота, називаються С3-рослинами, а рослини, в яких продуктом конденсації є оксалоацетат — С4- рослинами. До С4-рослин належать кукурудза, сорго, просо, цукрова тростина. Утворення C-4-карбонових кислот особливо важливе значення має для рослин, які ростуть у тропіках і субтропіках, замінюючи і доповнюючи в них цикл Кальвіна. Оптимальна температура фотосинтезу для С3-рослин—15—25 °С, а для С4-рослин — 45 °С. Тому в умовах високих температур досягається висока продуктивність фотосинтезу.
При досить інтенсивному освітленні і низькому вмісті оксиду вуглецю (IV) в атмосфері, коли процес фотосинтезу гальмується, в деяких рослин спостерігається короткочасне підвищене виділення оксиду вуглецю (IV). Виявилось, що утворюється він за рахунок розпаду гліколевої кислоти, що нагромаджується при фотосинтезі. Це явище називається фотодиханням. За цих умов в хлоропластах з участю ри- булозодифосфаткарбоксилази відбувається утворення фосфогліколевої кислоти, яка в результаті транспорту через мембрани хлоропластів гідролізується, а утворена гліколева кислота переходить з хлоропластів у зв’язані з ними структури — пероксисоми. Тут з участю гліколатоксидази вона перетворюється в гліоксилеву кислоту, яка використовується для синтезу вуглеводів і амінокислот гліцину, серину. Утворення сахарів з гліколевої кислоти називається гліколатним фотосинтезом.
Фотодихання є побічним шляхом фотосинтезу, який відбувається з використанням кисню і виділенням оксиду вуглецю (IV). Чим нижча концентрація вуглекислого газу і вища концентрація кисню, тим інтенсивніше відбувається реакція:
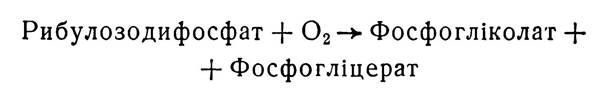
Фотодихання постачає досить важливі амінокислоти — гліцин і серин. Частина серину може перетворюватись у пероксисомах в гліцерат і включатися в цикл Кальвіна.
Тривалий час існувало уявлення про фотосинтез і фотодихання як два різних самостійних процеси, які відбуваються одночасно на світлі і в яких здійснюється взаємно протилежний обмін оксиду вуглецю (IV) і води.
На основі численних досліджень було встановлено, що ці два процеси взаємопов’язані між собою і зв’язуючою ланкою їх є фосфогліцеринова кислота. Залежно від умов вона використовується у двох напрямах — інтенсивне відновлення з участю НАД-Н+Н+ та енергії АТФ, внаслідок чого вуглекислий газ перетворюється в сахари, або ж дефос- форилювання фосфогліцеринової кислоти з наступним утворенням піровиноградної кислоти і ацетил-КоА, які включаються в цикл трикарбонових кислот, в результаті чого виділяється С02 (фотодихання). Інтенсивне виділення оксиду вуглецю (IV) відбувається на світлі переважно в складних фізіологічних умовах — при високих температурах, надмірному освітленні, пошкодженні фотосинтетичного апарату клітини.
При вивченні обміну вуглекислого газу в процесі фотосинтезу і дихання за різних фізіологічних умов було встановлено, що ці процеси взаємопов’язані і залежно від умов і функцій клітини забезпечують постійний синтез органічних речовин, необхідних для протікання метаболічних процесів.
Таким чином, абсолютна більшість темнових реакцій фотосинтезу відбувається з використанням енергії АТФ, однак, речовини, що утворюються в темновій фазі — вуглеводи, жири, білки — беруть участь в енергетичному обміні рослин і є важливим джерелом енергії. Першим продуктом фотосинтезу є фосфогліцеринова кислота, а кінцевим — фруктозо-6-фосфат, що використовується для синтезу вуглеводів і відновлення первинного акцептора оксиду вуглецю (IV)—рибулозо-1,5-дифосфату. Темнові реакції фотосинтезу надзвичайно складні і повністю не з’ясовані. Остаточне вивчення хімізму фотосинтезу дасть змогу успішно здійснити складні синтези органічних речовин за звичайних умов — при атмосферному тиску і низьких температурах. Значення цього важко переоцінити. В. І. Вернадський зазначав, що коли буде повністю розгадано таємниці хімізму фотосинтезу, стане можливим практичне використання невичерпних джерел оксиду вуглецю (IV), що міститься в карбонатних породах, та безпосереднє синтезування продуктів харчування.
Хемосинтез
Використання енергії сонячного випромінювання — важливий, але не єдиний спосіб синтезу органічних речовин з неорганічних. Поширеною є також інша форма автотрофної асиміляції — утворення органічних речовин з використанням енергії екзотермічних реакцій окислення різних речовин — сірководню, аміаку, азотистої кислоти, сполук металів (Mn, Fe) з нижчою валентністю. Цей шлях асиміляції називається хемосинтезом, він властивий багатьом бактеріям і мікроорганізмам, у яких відсутній зелений пігмент — хлорофіл. У результаті хемосинтезу з участю оксиду вуглецю (IV) та енергії окислення неорганічних сполук утворюються органічні речовини, які використовуються для забезпечення метаболічних процесів.
Процес хемосинтезу відкрив російський учений С. М. Виноградський, який докладно вивчив його особливості та встановив існування в природі автотрофних сірко-, залізо- та нітрифікуючих бактерій. На відміну від фотосинтезу, хемосинтез є аеробним процесом, тобто для його здійснення необхідна присутність кисню в навколишньому середовищі. Продукти окислення, як правило, виділяються в навколишнє середовище.
При дослідженні хімізму хемосинтезу з використанням міченого вуглекислого газу (14С02) було встановлено, що асиміляція його у хемосинтезуючих бактерій здійснюється з участю рибулозо-1,5-дифосфату. Внаслідок асиміляції оксиду вуглецю (IV), як і при фотосинтезі, утворюється первинний продукт — фосфогліцеринова кислота. Частина електронів, які виділяються внаслідок окислення неорганічних речовин, використовується для відновлення НАДФ+, а деякі — через систему цитохромів передаються на кисень. У процесі транспорту електронів на певних ділянках окислювального ланцюга відбувається синтез макроергічної сполуки — АТФ.
Досить важливу групу хемосинтезуючих організмів становлять нітрифікуючі бактерії, які забезпечують процеси нітрифікації в природі. Ці бактерії містяться в грунті і окислюють аміак, що утворюється в процесі гниття органічних решток, до азотної кислоти. Утворена азотна кислота реагує з мінеральними речовинами грунту, внаслідок чого утворюються нітрати — амонійні солі, які можуть використовуватись рослинами. У разі відсутності вологи та в певних специфічних умовах у грунті може нагромаджуватись досить велика кількість нітратів.
Процеси нітрифікації здійснюються двома видами бактерій— нітритними (Nitrosomonds) і нітратними (Nitrobakter). Перші здійснюють окислення аміаку, що утворюється в грунті внаслідок розпаду органічних речовин (гниття білків), до азотистої кислоти:
![]()
Енергія, яка виділяється внаслідок окислення аміаку, використовується для синтезу органічних сполук відновленням оксиду вуглецю (IV).
Утворена азотиста кислота окислюється нітратними бактеріями до азотної кислоти:
![]()
З рівнянь реакцій видно, що процес окислення аміаку є енергетично більш вигідним, ніж окислення азотистої кислоти.
Нітрифікуючі бактерії — яскравий приклад симбіозу нижчих організмів: бактерії, що окислюють азотисту кислоту, одержують її внаслідок життєдіяльності бактерій, які окислюють аміак.
Характерною особливістю нітрифікуючих бактерій є те, що вони можуть розвиватись при повній відсутності органічних речовин, які необхідні для розвитку інших організмів, проте, життєдіяльність їх неможлива без наявності бактерій і мікроорганізмів, які розкладають у грунті органічні рештки. Процеси нітрифікації здійснюються в природі досить широко і забезпечують нагромадження великої кількості нітратів. Таким чином, продукти життєдіяльності нітрифікуючих бактерій є важливим фактором підвищення родючості грунтів та забезпечення повноцінного мінерального живлення рослин, що є важливим фактором підвищення врожайності сільськогосподарських рослин.
У водоймах, де інтенсивно відбувається відмирання органічних решток рослинних і тваринних організмів і нагромаджується велика кількість сірководню, процес хемосинтезу здійснюється з участю сіркобактерій. Енергію, необхідну для синтезу органічних речовин, вони добувають внаслідок окислення сірководню:
![]()
Вільна сірка, що виділяється в результаті реакції, нагромаджується в безбарвних клітинах бактерій, і при нестачі сірководню окислюється до сірчаної кислоти, з якої потім можуть утворюватись доступні для рослин сульфати: 2S + 3О2+ 2H2О→2H2SО4+ 650 кДж
Енергія, що утворюється внаслідок окислення сірки, використовується для синтезу органічних речовин з участю вуглекислого газу.
Велика кількість сіркобактерій є в деяких зонах Чорного моря, де на глибині 200 м вода насичена сірководнем.
Хемосинтетичні бактерії, які окислюють сполуки заліза і марганцю нижчої валентності, також досить поширені в природі. Особливо велика кількість їх у водоймах. При цьому гідроксиди осідають на дно водойм і утворюють поклади болотної руди. Вважають, що велика кількість покладів залізомарганцевих руд є результатом життєдіяльності цих бактерій у далекі геологічні епохи.
Відома також група бактерій, здатних окислювати водень, метан та інші сполуки. Водневі бактерії постійно окислюють водень, який утворюється при анаеробному розкладі різних органічних речовин мікроорганізмами грунту.
![]()
Ці бактерії можуть розвиватись також на розчинах глюкози, тобто вони не є строго специфічними, як інші види бактерій.
Хемосинтетичні бактерії відіграють важливу роль, деякі з них мають народногосподарське значення. Сірчані бактерії очищають забруднені водойми, нітрифікуючі — затримують у грунті азот аміаку, що утворюється внаслідок гниття органічних решток. Хемосинтезуючі бактерії є типовими автотрофами, оскільки самостійно здійснюють синтез органічних речовин з неорганічних. Від фотосинтезуючих організмів вони відрізняються тим, що повністю не залежать від наявності джерела світла.
Значення фотосинтезу. Космічна роль рослин
Роль фотосинтезу в забезпеченні різноманітних метаболічних процесів, які протікають в біосфері і є основою життєдіяльності живих організмів, надзвичайно важлива. У процесі фотосинтезу неорганічні, позбавлені запасу хімічної енергії, інертні речовини — СО2, Н2О, мінеральні солі перетворюються в макроергічні органічні сполуки, які використовуються людиною і тваринним світом. Тому значення фотосинтезу насамперед полягає в тому, що він є першоджерелом утворення продуктів харчування для людини і тварин та найголовнішим процесом у живленні рослинних організмів. Усі інші процеси живлення лише стимулюють і поліпшують фотосинтетичну діяльність рослин, сприяють
кращому використанню продуктів фотосинтезу для різноманітних метаболічних процесів. З участю зелених рослин щороку синтезується велика кількість органічних речовин. У перерахунку на глюкозу це становить 450 млрд. тонн. Фотосинтез вважається найбільшим і найважливішим синтезом на Землі.
Завдяки фотосинтезу у природі нагромаджуються запаси енергії у вигляді різноманітних корисних копалин (вугілля, газу, нафти, торфу), які використовуються людиною для різноманітних потреб.
Для добування продуктів харчування людина також широко використовує природні ресурси — продукти фотосинтезу, а також культивує культурні високопродуктивні сільськогосподарські рослини. Площа орних земель, зайнятих посівами культурних рослин на нашій планеті, становить близько 2,9 млрд. гектарів, з яких щороку люди одержують 10 млрд. тонн органічної біомаси, або 600—700 млн. тонн органічних речовин харчових продуктів. Підраховано, що продукти фотосинтезу становлять 96 % речовин, які людина використовує в побуті, техніці, для харчових потреб. Для здійснення такого широкомасштабного синтезу органічних речовин рослинні організми використовують велику кількість вуглекислого газу. Сільськогосподарські рослини в процесі фотосинтезу засвоюють з повітря за добу 120— 240 кг СО2 на 1 гектар посівів. При цьому поверхня зелених рослин площею 1 м2 асимілює за 1 год 1,5—2,5 г СО2 і синтезує 1—1,5 г вуглеводів. Якщо врахувати, що в 1 м3 повітря міститься близько 0,5 г СО2, то для синтезу 1 г вуглеводів рослина повинна переробити 3 м3 повітря. Щороку для потреб фотосинтезу рослинами використовується близько 10 % всього вуглекислого газу, що є в атмосфері. Ця величина становить 0,4·10 11 тонн, з них 7г припадає на зелені рослини суші, а решта — на рослини морів і океанів.
Вміст асимільованого рослинами вуглекислого газу в атмосфері компенсується промисловими викидами, вулканічною діяльністю, а також різноманітними окислювальними процесами, як суто хімічними (горіння), так і біохімічними (дихання, біологічне окислення). Стає зрозумілою санітарна роль зелених насаджень, особливо в великих промислових містах, де в результаті діяльності підприємств, спалювання вугілля та інших видів палива в атмосферу потрапляє велика кількість оксиду вуглецю (IV), внаслідок чого вміст його може в багато разів перевищувати нормальний рівень (0,03%). Тому зелені рослини відіграють
важливу роль у підтримуванні балансу вуглекислого газу в атмосфері нашої планети, що має надзвичайно важливе значення. Адже завдяки вмісту вуглекислого газу та водяної пари регулюється температура земної поверхні. Обчислено, що збільшення вмісту СО2 в атмосфері в 2—2,5 рази привело б до підвищення температури поверхні на 8 °С.
А. Л. Курсанов встановив, що вуглекислий газ може засвоюватись рослинами не лише з повітря, а і з грунту за допомогою кореневої системи. Основним джерелом вуглекислого газу є органічні рештки, які розкладаються мікроорганізмами, та процеси дихання кореневої системи рослин. Методом мічених атомів встановлено, що кількість оксиду вуглецю (IV), засвоєного завдяки кореневій системі, може становити близько 5 % загальної кількості її, що ввібрана рослиною. Засвоєння оксиду вуглецю (IV) з грунту здійснюється з участю ферментів карбоксилаз, які забезпечують процеси карбоксилювання піровиноградної кислоти, утвореної внаслідок розпаду глюкози і перетворення її в щавлевооцтову кислоту. Утворена щавлевооцтова кислота є основним субстратом циклу трикарбонових кислот, який є зв’язуючою ланкою між різними видами обміну речовин — вуглеводного, ліпідного, білкового.
Оскільки фотосинтез забезпечує одержання високоякісних врожаїв сільськогосподарських культур, то велике значення має широке застосування методів і агротехнічних прийомів, які б сприяли підвищенню ефективності фотосинтезу і збільшенню коефіцієнта корисної його дії.
Доведено, що фотосинтез, кореневе живлення та водообмін перебувають в тісному взаємозв’язку, тому в науково-дослідних закладах нашої країни постійно проводиться дослідження по вивченню впливу світлового режиму, густоти насадження, способів посіву та підживлення рослин на ефективність фотосинтезу.
Оволодіння таємницями фотосинтезу, розробка та впровадження заходів щодо підвищення коефіцієнта корисної його дії — досить важлива проблема, над якою працюють учені-біологи, фізіологи, біофізики, біохіміки. Оволодіння методами ефективного керування цим процесом та штучного його відтворення дасть можливість набагато підвищити ефективність сільського господарства, збільшити виробництво продуктів харчування для населення. Оскільки проблема харчових ресурсів є однією з найважливіших і найгостріших для людства, то успішне розв’язання її є основним завданням біологічної і сільськогосподарської науки.
Важливу роль відіграють зелені рослини як продуценти кисню, необхідного для всіх живих організмів. Виділення кисню в процесі фотосинтезу мало велике значення в еволюції життя на Землі. Первинна, відновна атмосфера, поступово збагачуючись киснем, перетворилась у кисневмісну окислювальну оболонку. А це в свою чергу забезпечило можливість існування життя. Зелені рослини виділяють вільний молекулярний кисень, який використовується людиною і тваринами для забезпечення процесів життєдіяльності. Важливу роль у цьому процесі відіграють не тільки рослини суші, а й численні водорості, які населяють моря і океани планети. За один рік водорості виділяють 3,6·1011 тонн кисню, що становить 90 % всього кисню, який потрапляє в атмосферу з поверхні планети. Тому саме водорості вважають основним джерелом кисню, саме вони створюють ту частину атмосфери, яка необхідна для існування живого. Наше життя нерозривно пов’язане з «диханням океану» і безперервною фотосинтетичною діяльністю водоростей.
За добу людина в середньому використовує близько 500 л кисню. А за рік все населення планети споживає близько 900 млрд. кубічних метрів кисню. Значна кількість кисню використовується тваринним світом та промисловістю. В зв’язку з цим стає зрозумілим, що без постійного поновлення запасів кисню життя було б неможливим. Таким чином, життя людини і тварин перебуває в подвійній залежності від фотосинтетичної діяльності рослин. Рослини продукують органічні речовини, які використовуються гетеротрофними організмами і, як побічний продукт, виділяють необхідний для життя кисень. У цьому полягає важлива космічна роль рослин, що в свій час відзначав К. Д. Тімірязєв. Потенціал продуктивності рослин досить високий, однак від усієї кількості органічних речовин, які синтезуються зеленими рослинами, ми використовуємо лише незначну кількість у вигляді рослинної їжі або у видозміненому вигляді, як продукти тваринного походження.
Проблема найкращого використання фотосинтетичної продуктивності рослин — одна з найважливіших у діяльності людини. Успішне її розв’язання тісно пов’язане з охороною навколишнього середовища, збільшенням зелених насаджень, економним витрачанням сировини і матеріалів.