БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
27. МЕХАНИЗМЫ ВИДООБРАЗОВАНИЯ
27.3. Отбор
Отбор можно рассматривать как механизм, действующий на двух связанных между собой уровнях — на уровне организма и на уровне аллелей.
Отбор — это процесс, в результате которого организмы по своей морфологии, физиологии и поведению лучше адаптированные к данной среде, выживают и размножаются, а менее адаптированные либо гибнут, либо не могут оставить потомство. Первые передают свои благоприятные признаки следующему поколению, а вторые нет. Отбор зависит от существования в популяции фенотипической изменчивости и составляет часть механизма, обеспечивающего адаптированность вида к своей среде.
При увеличении численности популяции некоторые факторы среды, такие как пища — для животных — и свет — для растений — становятся лимитирующими. Это приводит к конкуренции за ресурсы между членами популяции. Организмы, признаки которых дают им преимущество в конкуренции, легче овладевают этими ресурсами, выживают и оставляют потомство. Организмы, лишенные таких признаков, оказываются в невыгодном положении и могут погибнуть, прежде чем успеют произвести потомство. Совместное действие лимитирующих факторов среды и численности популяции создает давление отбора, интенсивность которого может быть различной.
Таким образом, отбор — это процесс, определяющий, какие аллели будут переданы следующему поколению благодаря относительным преимуществам, которые они доставляют, экспрессируясь в фенотипе. Следовательно, давление отбора можно рассматривать как способ увеличения или уменьшения распространенности какого-либо аллеля в генофонде, и эти изменения частоты аллелей могут вести к эволюционному изменению.
Крупные изменения в генотипе возникают в результате распространения в генофонде мутантных аллелей. Степень отбора и его скорость зависят от характера мутантного аллеля и силы его влияния на данный фенотипический признак. Если аллель доминантен, то он проявляется в фенотипе чаще, и отбор начинает оказывать на него положительное или отрицательное давление быстрее. Если же аллель рецессивен и не проявляется в гетерозиготном состоянии, как это обычно для большинства мутаций, то он не подвергается отбору до тех пор, пока не появится в гомозиготном состоянии. Вероятность быстрого появления таких рецессивных гомозигот невелика, и новый аллель может исчезнуть из генофонда, прежде чем они возникнут. Рецессивный аллель, неблагоприятный в данной среде, может сохраниться в популяции и дождаться таких изменений среды, при которых он будет обладать преимуществом. Вероятно, эти эффекты проявятся сначала у гетерозиготы, и отбор будет благоприятствовать ее распространению в популяции, как в случае серповидноклеточной анемии.
Рецессивный мутантный аллель может быстро распространиться в популяции, если его локус в хромосоме сцеплен (находится в тесной близости) с локусом какого-либо доминантного аллеля, имеющего важное функциональное значение и подвергающегося сильному положительному отбору. При таком сцеплении шансы мутантного аллеля соединиться с другим мутантным аллелем и оказаться в гомозиготном состоянии сильно возрастают (рис. 27.1).

Рис. 27.1. Ускоренное распространение в популяции мутантного аллеля (а), сцепленного с доминантным аллелем (В), которому сильно благоприятствует отбор.
Влияние каждого данного мутантного аллеля может варьировать. Мутации, затрагивающие аллели, контролирующие важные функции, вероятнее всего окажутся летальными и будут немедленно элиминированы из популяции. Эволюционные изменения происходят обычно в результате постепенного появления мутантных аллелей, производящих небольшие прогрессивные изменения в фенотипических признаках.
Существуют три типа отбора, происходящего в природных и искусственных популяциях: стабилизирующий, направленный и дизруптивный. Их легче всего объяснить, построив кривые нормального распределения для непрерывной фенотипической изменчивости, наблюдаемой в природных популяциях (рис. 27.2).
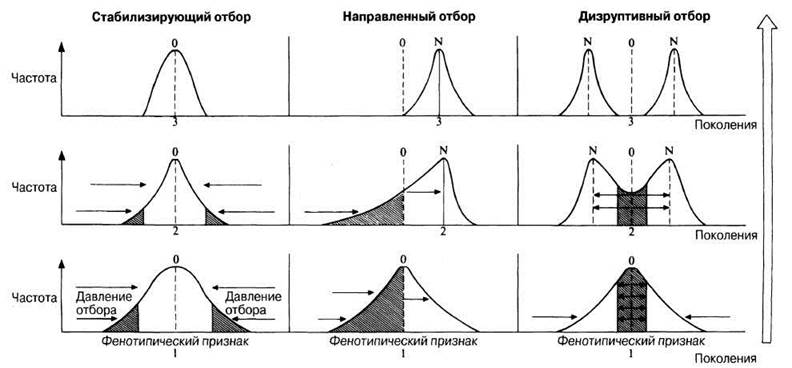
Рис. 27.2. Три типа отбора, действующие в популяциях. 0 — первоначальное положение точки соответствия между оптимальным фенотипом и оптимальными условиями среды; N — новое положение точки соответствия. Особи, признаки которых попадают в заштрихованные участки, оказываются в невыгодном положении и элиминируются отбором. Цифрами 1—3 указана последовательность поколений.
27.3.1. Стабилизирующий отбор
Стабилизирующий отбор происходит в тех случаях, когда фенотипические признаки соответствуют оптимальным условиям среды и конкуренция относительно слабая. Такой отбор действует во всех популяциях, элиминируя из них особей с крайними отклонениями признаков. Например, существует некая оптимальная длина крыла для хищной птицы определенных размеров и определенным образом жизни в данной среде. Стабилизирующий отбор, действуя благодаря дифференциальному размножению, будет элиминировать тех птиц, у которых размах крыльев больше или меньше оптимального.
Карн и Пенроуз (Каш, Penrose) изучали корреляцию между массой новорожденных и постнатальной смертностью у 13 730 младенцев, родившихся в Лондоне в период с 1935 по 1946 гг. Из них 614 родились мертвыми или умерли в первый месяц после рождения. Как показывает рис. 27.3, оптимальная для новорожденных масса равна примерно 3,6 кг. Младенцы с большей или меньшей массой оказывались в смысле отбора в невыгодном положении, и смертность среди них была несколько выше. На основе этих данных можно вычислить интенсивность давления отбора.

Рис. 27.3. Зависимость между массой тела новорожденных и их выживанием. (По М. Каrn, L. S. Penrose, Ann. Eugen., 1951, 16, 147-164).
Если 614 младенцев умерли при рождении или в первый месяц жизни, то смертность составляет 4,5%. Даже среди младенцев с оптимальной массой смертность составляла 1,8%. Следовательно, давление отбора на массу при рождении, равную 3,6 кг, составляет 4,5% — 1,8% = 2,7%, или 0,027. При массе 1,8 кг смертность была равна 34%, что соответствует давлению отбора примерно 30%, или 0,3. Следует указать, однако, что в результате успехов, достигнутых педиатрией после 1946 г., постнатальная смертность значительно снизилась.
Стабилизирующий отбор не способствует эволюционному изменению, а поддерживает фенотипическую стабильность популяции из поколения в поколение.
27.3.2. Направленный отбор
Эта форма отбора возникает в ответ на постепенные изменения условий среды. Направленный отбор влияет на диапазон фенотипов, существующих в данной популяции, и оказывает селективное давление, сдвигающее средний фенотип в ту или другую сторону. После того как будет достигнуто оптимальное соответствие между средним фенотипом и новыми условиями среды, вступает в действие стабилизирующий отбор.
Направленный отбор приводит к эволюционному изменению, оказывая на популяцию такое давление, которое благоприятствует увеличению в ней частоты новых аллелей. Направленный отбор лежит в основе искусственного отбора, при котором избирательное скрещивание особей, обладающих желательными фенотипическими признаками, повышает частоту этих признаков в популяции (см. разд. 27.4). В ряде экспериментов Фалконер (Falkoner) выбирал из популяции шестинедельных мышей самых тяжелых особей и давал им спариваться друг с другом. То же самое он проделывал с самыми легкими мышами. Такое избирательное скрещивание по признаку массы тела привело к созданию двух популяций, в одной из которых масса возрастала, а в другой уменьшалась (рис. 27.4). После прекращения селекции ни та, ни другая группы не вернулись к первоначальной массе (примерно 22 г). Это показывает, что искусственный отбор по фенотипическим признакам привел к некоторому генотипическому отбору и к частичной утрате обеими популяциями каких-то аллелей. В настоящее время непосредственно изучено много примеров классического направленного отбора; они описаны в разд. 27.5.
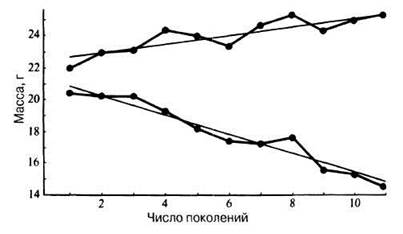
Рис. 27.4. Изменения массы тела в последовательных поколениях двух популяций мышей, подвергавшихся отбору по этому признаку. (По D. S. Falconer, J. Genetics, 1953, 51, 470-501.)
27.3.3. Дизруптивный отбор
Это, вероятно, самая редкая форма отбора, но она может играть очень важную роль в возникновении эволюционных изменений. Колебания условий в данной среде, связанные, например, со сменой времен года и климатическими факторами, могут благоприятствовать наличию в данной популяции не одного, а двух или нескольких фенотипов. Давления отбора, действующие в самой популяции, могут привести к отклонению фенотипов от среднего для данной популяции к обоим крайним типам. В результате популяция окажется разделенной на две субпопуляции. Если поток генов между этими двумя субпопуляциями нарушится, то каждая из них может дать начало новому виду. В некоторых случаях такая форма отбора приводит к появлению в одной популяции нескольких различных фенотипов, или к полиморфизму (от греч. polýmorphos — многообразный), который будет рассмотрен в разд. 27.5.1. В пределах одного вида популяции с различными фенотипами, или экотипы, могут быть адаптированы к определенным условиям среды (разд. 27.6.2). Если вид занимает очень обширный географический ареал, то у населяющих этот ареал популяций могут наблюдаться локальные изменения фенотипических признаков, в результате чего они становятся промежуточными между популяциями краевых областей. Такой непрерывный градиент постепенно изменяющихся признаков на протяжении географического ареала представляет собой обычно фенотипическую реакцию на изменение климатических и(или) эдафических (почвенных) факторов и известен под названием клины (разд. 27.6.3).
27.3.4. Интенсивность давления отбора
Интенсивность давления отбора в данной популяции варьирует в пространстве и во времени, что может быть связано с изменениями внешних или внутренних факторов. К внешним факторам относятся, в частности, увеличение численности хищников или патогенных организмов и конкуренция с другими видами (межвидовая конкуренция) за пищу и места для размножения (у животных) или за свет, воду и минеральные вещества (у растений). Изменения климата или состояния местообитания могут привести к созданию новых давлений отбора. Внутренние факторы, например быстрое повышение численности популяции, нередко приводят к усилению конкуренции за ресурсы (внутривидовая конкуренция). С увеличением популяции данного организма возрастает и численность его паразитов и хищников; кроме того, в многочисленной популяции облегчается передача паразитов и болезней от одной особи к другой. Все эти факторы могут изменять не только интенсивность давления отбора, но и его направление. Отбор благоприятствует «новым» фенотипам (и генотипам), а плохо адаптированные особи элиминируются. В первую очередь при этом устраняются особи, обладающие неадаптивными крайними фенотипами.
Одним из результатов возросшего давления отбора может быть специализация организмов к определенному образу жизни или более узкому диапазону условий среды. Не исключено, что это окажется неблагоприятным для дальнейшей судьбы вида. Увеличение единообразия вида и его зависимости от каких-либо определенных условий повышает вероятность его вымирания в случае изменения этих условий. В палеонтологической летописи встречается немало вымерших организмов, которые были крайне своеобразны и чересчур специализированы.
27.2. Как сведения о давлении отбора в образе жизни определенного паразита могут помочь искоренить его?
Из всего сказанного можно видеть, что повышенное давление отбора — это консервативный механизм, сохраняющий фенотип, лучше всего адаптированный к данным условиям среды (оптимальный фенотип).
Ослабление интенсивности отбора обычно оказывает противоположное действие. Оно может наступить при отсутствии хищников, патогенных организмов, паразитов и конкурентов или же при улучшении условий среды. Так обстоит дело при проникновении вида в новую для него среду. Как полагают, именно такая ситуация благоприятствовала созданию видового разнообразия вьюрков на Галапагосских островах.